Information Links
Related Conferences
Previous Issues Volume 7, Issue 2 - 2024
Elimination/Neutralization of COVID-19 Vaccine-Produced Spike Protein: Scoping Review
Steven Petrosino1-3*, Nyona Matende4
Pennsylvania State University, USA
LaSalle University, USA,
Walden University, USA
Kenya Medical Research Institute (KEMRI)
*Corresponding Author: Steven Petrosino, Pennsylvania State University, LaSalle University, and Walden University, USA; Email: [email protected]
Received Date: January 23, 2024
Publication Date: April 12, 2024
Citation: Petrosino S, et al. (2024). Elimination/Neutralization of COVID-19 Vaccine-Produced Spike Protein: Scoping Review. Mathews J Nutr Diet. 7(1):34.
Citation: Petrosino S, et al. © (2024)
ABSTRACT
This review investigates the long-term sequelae of SARS-CoV-2 spike protein persistence following COVID-19 vaccination, its immunopathology, its impact on host immunity, and potential strategies for its neutralization or elimination. The urgency with which vaccines were required at the height of the COVID-19 pandemic led to a failure to consider the adverse long-term effects of the spike-protein in the mRNA vaccines developed by leading manufacturers. The present review study was completed in eight-stage PRISMAScR procedures. A total of 420 studies were identified for potential inclusion, with a final 40 meeting the criteria for inclusion in the spike protein neutralization, elimination, or removal assessment. Numerous biological molecules, synthetic and natural compounds were found to possess promising spike protein neutralization, elimination or removal properties {Antibodies (monoclonal antibodies at 25%, neutralizing antibodies at 12.5%, egg yolk antibodies at 7.5%, non-neutralizing enzymes at 2.5%); Nanomaterials (liposomes at 5.0%, Encapsulin nanoparticles at 2.5%, polymeric NPs at 2.5%, metalliv NPs at 2.5%); Natural compounds (polyphenols or green tea at 7.5%, Urea at 2.5%); Engineered proteins (DARPins - FSR16m and FSR22 at 2.6%, Nattokinase at 2.5%); Radiation (Low dose radiation therapy at 2.5%); Equipment (Hemopurifier equipment utilizing lectin-affinity plasmapheresis cartridge at 2.5%)}. The supplemental section provides additional compounds reported in grey literature with independently verified potential to neutralize, eliminate or remove spike protein, including vitamin C, vitamin D, Omega 3 (fish oil), curcumin/turmeric, ecinacea, resveratrol, dandelion, and calendula. The supplemental compounds must be used with the direction of a healthcare provider.
INTRODUCTION
The worst of the COVID-19 pandemic may be over but residual effects of the virus may be far more deadly than most understand. The long-term sequelae of the viral infection and of the pandemic response measures, including COVID vaccination, treatment with remdesivir, and the widespread in-hospital use of intubation and mechanical ventilation have created a new group of patients with pressing healthcare needs that may not be completely understood [1]. Emerging from the Wuhan region of central China in late 2019, the novel Coronavirus (SARS-CoV-2) created fear and anxiety and ravaged the world as a global pandemic with far reaching catastrophic implications. Like other viruses, specific viral proteins are critical for the ability of the novel coronavirus to both replicate and invade and fuse with human cells. Spike proteins adorning the outer shell of the SARS-CoV-2 virus are critical in the binding of the viral particle to the human host cell membrane, allowing the viral RNA to enter into host cells, hijack the machinery of the compromised cell nucleus and facilitate viral replication [2]. Utilizing the Receptor Binding Domain (RBD), SARS-CoV-2 spike proteins bind to Angiotensin Converting Enzyme-2 (ACE2) expressing cells in the nasal passages, lungs, and upper respiratory tract to facilitate viral infection [3].
The interaction between the RBD of the viral spike glycoprotein with the somatic ACE2 receptor in the Protease Domain (PD) of host human cells is crucial for the ability of the novel coronavirus to infect humans and, hence, precipitate a global pandemic [4]. In less than a year into the pandemic, epidemiologists and other medical scientists had sufficiently understood the virus to recommend measures capable of safeguarding the health of those at greatest risk of adverse outcomes (the elderly, the immunocompromised and those suffering from comorbidities) and to limit the impact on livelihoods, particularly among people with a low risk of serious COVID-19 illness (primarily the young and healthy) [5]. However, the sense of urgency created by the mainstream press, leaders of both governmental parties, major government agencies, and uncensored vocal medical professionals led to the swift development, rapid but inadequate evaluation, and expeditious Emergency Use Authorization (EUA) of vaccines specifically designated only for “emergency use” in less than a year. This was termed “Operation Warp Speed” by the current administration in early 2020. The average gestation period for the development of a new vaccine is 10 to 11 years [6]. The lengthy process taken in the course of vaccine development is occasioned by the need for a “proof of concept” involving an exploratory stage, a pre-clinical stage (phase I), followed by phase II and phase III clinical trials, and subsequent regulatory review and approval [7]. For COVID-19 vaccines to be produced in less than a year, it was obvious to researchers that they were required to be enormously accelerated. In one study, 65% of the respondents agreed that announcement of vaccine-release in less than a year following the initial virus outbreak implied that the vaccines were rushed [8]. The two major COVID-19 vaccines were mRNA-based (messenger RNA), utilizing only the full-length spike protein part of the virus. Researchers uniformly neglected other viral protein targets, such as the envelope or nucleocapsid. The uniqueness of the spike protein and location on the surface of the virus made it exceptionally distinctive for immune memory cell targeting and presumed safe since it did not contain other viral particles [9]. Pfizer and Moderna pharmaceutical companies, the formulators of these spike protein vaccines did not consider that the spike protein itself might possess toxic characteristics.
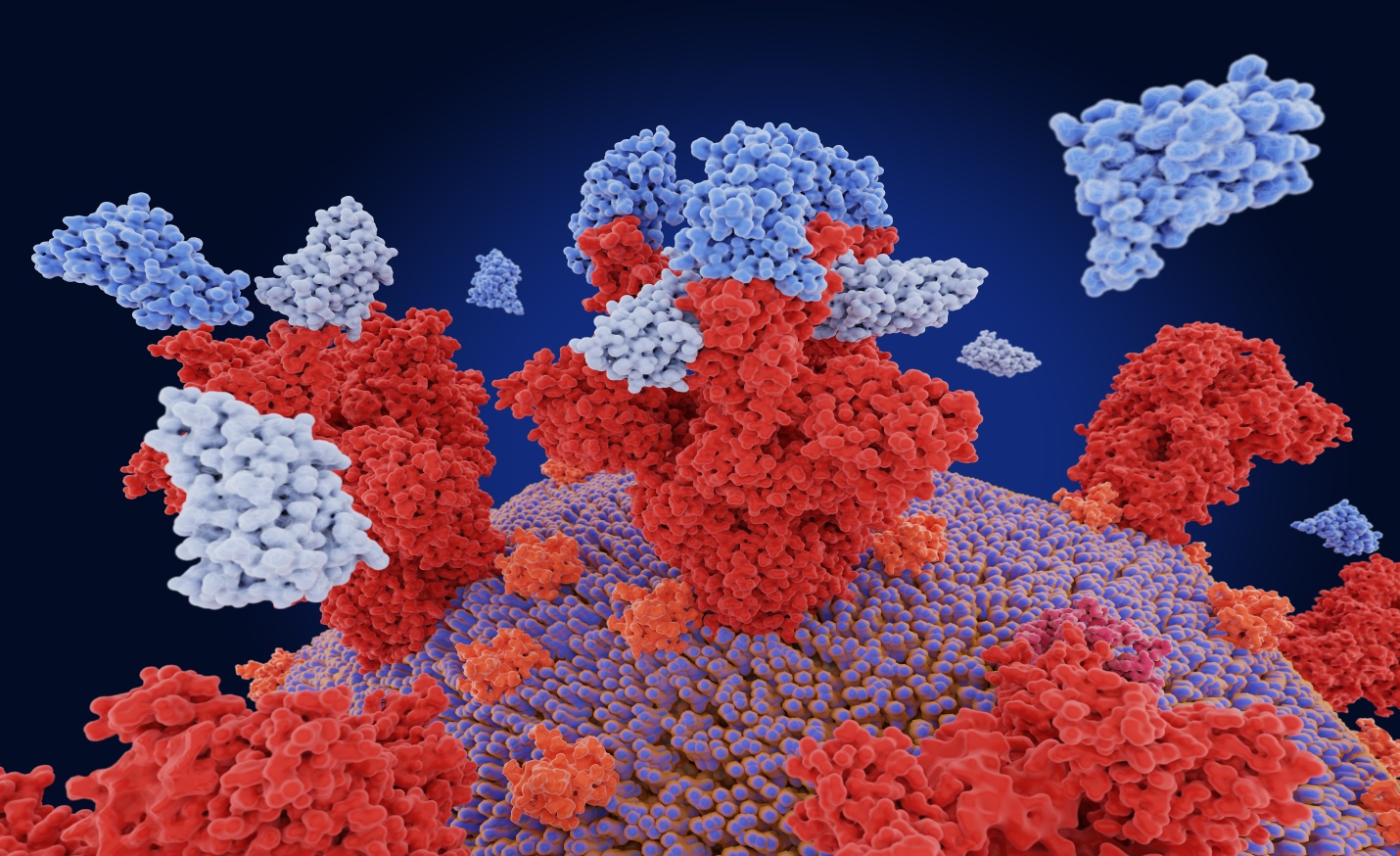
Figure 1: COVID-19 spike protein binding to host ACE2 cells.
Figure 1: Spike Protein (S), found on the surface of COVID-19 virus cells binds to pericytes, which line the small blood vessels of the heart. This binding triggers a cascade of changes that disrupt normal cell function. Receptor Binding Domain (RBD) or the uncoated loop sticking out of the top of the spike protein provides one of three sections of the spike that bind to receptors on human cells. When the RBD lifts above the glycan cloud, two glycans swoop in to lock it into place, enhancing its ability to bind to human cell receptors. One of the functional units binds to ACE2 receptor triggering the uptake of the virus particle and eventually membrane fusion, allowing the virus to enter and infect host cells.
The vaccines were predicated on the introduction of mRNA in muscle cells of the deltoid to hijack the cellular replication machinery of these cells to induce their production of copies of spike protein. The injected mRNA (which is inherently fragile and unstable) was transiently enveloped in a protective polyethylene glycol (PEG) casing designed to degrade within a few days after delivering its fragile payload [10]. The hurried development of the COVID-19 vaccine, under the auspices of Operation Warp Speed, established that these mRNA vaccines were able to induce a robust immune response against SARS-CoV-2, but this whirlwind developmental pace overlooked (or simply ignored) the fact that the molecular mode of action (MOA) was not completely understood, as well as the potential adverse events associated with this novel therapy [11].
Several studies have subsequently demonstrated that full-length spike protein, unbound by antibodies, continues to circulate for an extended period post-vaccination, occasioning significant morbidity and mortality [11].
A peer-reviewed study in mice found that the spike protein can cross the blood-brain barrier. The study showed that circulating spike can navigate to the brain, bodily organs and other tissues, and it is quite possible that blood tests looking for its presence will be negative while it sequesters and occultly persists in bodily organs, the lining of blood vessels, and other bodily tissues [12].
In a biodistribution study conducted in Japan on Pfizer SARS-CoV-2 mRNA Vaccine (BNT162b2-encapsulated LNP) vaccine found that 48 hours post-vaccination, vaccine particles had migrated to various tissues throughout the body and did not stay at the deltoid muscle injection site, with high concentrations found at the liver, bone marrow, and ovaries [13].
COVID-19 Vaccination and Spike Protein Persistence
Recently, research has suggested that one primary difference between vaccine mediated immunity and immunity acquired from natural infection is the mRNA vaccine’s ability to pass the first line of defense of innate immunity against infection, the mucous cells of the nasal pharynx and tonsils which are triggered by natural infection. The mRNA vaccines, like other vaccines, stimulate an adaptive immune response, which can subsequently target the virus in these tissues. The adaptive immune response is an immune response that is specific to the attacking pathogen and is also called “acquired immunity” or “specific immunity”. The adaptive immune response utilizes B cells which produce antibodies against the specific pathogen, and T cells which attack and destroy infected cells. The adaptive immune system is slower to respond than the innate immune system, but it is more specific and can provide long-lasting protection against infection. The mRNA vaccines are typically administered through the intramuscular route. After being injected into the muscle (usually the deltoid muscle), the vaccine is taken up by immune cells and transported through the lymphatic system to the lymph nodes, where it can stimulate an immune response [14]. However, it is possible for the vaccine to be inadvertently injected into a blood vessel. This is known as intravascular injection and can result in the vaccine being delivered directly into the bloodstream rather than the muscle. Studies have estimated that this may happen in about 1 in 800 injections, although the actual rate may vary depending on the technique used for injection. The phenomenon may result in the vaccine being delivered directly to the cardiac muscle, which may be responsible for the increased incidence of myocarditis which has been observed following COVID-19 mRNA vaccination [14]. A recent report by the Australian Health Authority demonstrated that the mRNA vaccine lipid nanoparticles are distributed throughout the body, including concentrations in the lymph nodes, spleen and bone marrow, and this report also found that the spike protein was present in the blood for up to 14 days following mRNA vaccination (T. It has been reported that certain antibodies in the blood of COVID-19 infected patients appear to change the shape of the spike protein to make it more likely to bind to cells, while other studies showed that the spike protein by itself (without being part of the corona virus) can damage endothelial cells and disrupt the blood-brain barrier [14]. These findings may be even more relevant to the pathogenesis of long-COVID syndrome that may affect as many as 50% of those infected with SARS-CoV-2. The results suggest that the spike protein generated by the vaccine may also have toxic potential. Yonker et al established a “markedly elevated levels of full-length spike protein, unbound by antibodies, were detected in the plasma of individuals with postvaccine myocarditis, whereas no free spike was detected in asymptomatic vaccinated control subjects (P<0.0001)” [15].
Needle aspiration, a technique used to ensure that the needle is not in a blood vessel before injecting the vaccine, may reduce the risk of intravascular injection, and may potentially reduce the risk of myocarditis, however both the WHO and CDC stopped recommendations for needle aspiration in 2016, and the practice is less common today. A limited impromptu survey of several local pharmacies and medical centers offering the COVID-19 vaccine by this author (SPP) found that no administering HCPs were using the aspiration technique. According to the CDC, “aspiration before injection of vaccines or toxoids is not necessary for the recommended injection sites” [16]. This is because no large blood vessels are present in those locations, findings corroborated in another study [17].
Mechanisms of Spike Protein Harm
As previously noted, the role of the spike protein (SP) in SARS-CoV-2 viral infection has been thought to be limited to the entry of the virus into host cells through mediation of ACE2 receptor-binding and fusion. However, recent findings show the possibility of direct SP involvement in certain COVID-19 pathological presentations following natural infection. These adverse effects may also present following COVID-19 vaccination. The SARS-CoV-2 vaccination adverse effects (AEs) have been identified, with 1.5 million adverse events and 34,000 COVID deaths catalogued in the US Vaccine Adverse Events Reporting System (VAERS) as of this writing [18]. Numerous explanations for these events have been postulated, however the precise incidence of these events cannot be accurately determined as VAERS captures only between 1% and 10% of these vaccine-associated adverse events according to some experts [19,20]. Some of the speculations surrounding these events involve the proinflammatory action of the lipid nanoparticles used for the delivery of mRNA (i.e., the vaccine formulation), as well as theories concerning the unique nature, expression pattern, binding profile, and proinflammatory effects of the host immune system-produced antigens to the spike protein and/or its subunits/peptide fragments in human tissues or organs [11]. The lipid nanoparticle surrounding the mRNA of the COVID vaccine is composed of polyethylene glycol (PEG). It has been widely assumed that PEG would lack antigenicity or immunogenicity. However, with increased popularization of PEGylation technique in medicines, laxatives, and even bowel preps for colonoscopy, anti-PEG antibodies have been detected in not only patients treated with PEGylated therapeutics but also PEGylated drug treatment-naïve individuals with a prevalence of up to 72% [21]. These authors concluded that “the widespread prevalence of pre-existing anti-PEG Ab, coupled with high Ab levels in a subset of the population, underscores the potential importance of screening patients for anti-PEG Ab levels prior to administration of therapeutics containing PEG”. Some other studies have directly associated COVID-19 vaccination with worsened lipid toxicity, resulting in subsequent organ damage [22]. According to Avolio et al., (2021), the action of spike protein through a cluster of CD147 differentiation resulted in detrimental effects on human cardiac pericytes [22]. Spike proteins have also been shown to precipitate cardiovascular disease even in the absence of viral infection [23]. In another study involving immune-profiling of young individuals vaccinated against SARS-CoV-2, it was established that most of the respondents experienced myocarditis within the first week of vaccination [15]. In fact numerous adverse effects (AEs) have been reported following COVID-19 vaccination, including Bell’s palsy, cerebral venous sinus thrombosis, Guillain–Barré syndrome, myocarditis/pericarditis (mostly in younger subjects, with a male predominance), thrombosis with thrombocytopenia syndrome, pulmonary embolism, stroke, lymphadenopathy, appendicitis, herpes zoster reactivation, neurological complications, and autoimmunity (e.g., autoimmune hepatitis and autoimmune peripheral neuropathies [25-28]. The widespread prevalence of COVID-19 vaccination in the United States (At least 270,129,677 people or 81% of the population have received at least one dose at the time of this writing, suggests that the potential deleterious health impact of COVID-19 vaccination could be much worse compared to that of actual infection [29]. Numerous studies have indicated that the spike protein of SARS CoV 2 alone, in the notable absence of the whole COVID-19 virus, can have toxic effects. The following are some notable examples of studies relevant to the finding.
A study published in Neurobiology of Disease in October 2020 found that the spike protein promotes loss of blood-brain barrier integrity and triggers an inflammatory response in brain endothelial cells [30]. Another study published in Vaccines noted that the pathogenicity of the spike protein is a relevant concern for COVID 19 vaccines since the mRNA vaccines are designed to instruct human cells to produce the spike protein of the coronavirus [31]. It was also showed that spike protein alone can damage vascular endothelial cells in one study published in Circulation Research [32]. Bozkurt, Kamat, and Hotez (2021) offered an interesting perspective in a study published in Journal Circulation and which found that mRNA COVID 19 vaccines can cause myocarditis, suggesting that this might be due to individual immune responses to either vaccine mRNA or the vaccine-induced spike protein [33].
It is interesting that despite its commitment to full transparency, the FDA, on behalf of Pfizer, attempted to hide the full scope of the Pfizer-BioNTech COVID-19 vaccine adverse events for 75 years, prior to this attempted obfuscation being overturned in court [34]. It is therefore essential that measures are taken to fully address the adverse impacts of SARS-CoV-2 spike protein.
Study Objectives
The objective of the present study is to establish approaches and means through which SARS-CoV-2 spike protein can be neutralized or eliminated from the body and, therefore, prevent or mitigate some or all the deleterious effects of COVID-19 vaccination.
METHODS
A scoping review approach is found to be the most appropriate for the present investigation given the complexity, novelty, and broad nature of potential solutions to the identified problem. The intention is to identify the possible mechanisms, processes, compounds or formulations that can be used to eliminate residual spike protein following COVID-19 vaccination. The study follows the preferred reporting items for systematic reviews and meta-analysis extension for scoping reviews (PRISMAScR) procedures [35]. The present review was completed in eight stages: identification of appropriate sources; development of search terms; selection of sources; charting of retrieved data; definition of data items; critical appraisal of categories in relation to evidence; collating results; and provision of summaries for main findings for various categories.
Sources
Several reference databases were searched for literature published following the advent of the COVID-19 pandemic, with key ones including PubMed, EMBASE, and PubMed Central (PMC). Additional sources were searched through the Google Scholar search engine. Due to the sensitive nature of the topic (where partisan interests may affect the publication of scientific evidence on adverse impacts of vaccination) opinion from non-peer reviewed articles was considered if consistent with general hypothesis. However, the findings from the non-academic/peer-reviewed sources were separately analyzed in a supplemental section.
Search Terms
The novel nature of the research interest was taken into consideration during the development of a search strategy. Using a brief preliminary research, the authors created diverse search terms to capture the diverse nature of approaches/compounds that might be useful in the neutralization or elimination of COVID-19 spike protein. Some of the keywords noted in the preliminary research include antibodies (monoclonal and neutralizing antibodies, egg yolk antibodies), natural compounds (such as polyphenols), antioxidants, and COVID-19 antivirals). The table below (Table 1) summarizes the search strategy implemented in each of the utilized databases.
Table 1: Search Strategy.
|
Tier 1 Keywords: Spike Protein, COVID-19 Spike protein, Coronavirus Spike Protein, COVID-19 Vaccine Spike Protein Tier 2 Keywords: inhibitors, antibodies, nanomaterials, polyphenols, antioxidants, agonists, natural compounds, egg yolk antibodies, monoclonal antibodies, neutralizing antibodies. Tier 3 Keywords: Neutralization, removal, elimination, treatment. *In conducting the searches, keywords from the three tiers were combined using Boolean operators. |
Table 1: Tiers 1,2, and 3 identified broad categories of keywords, which were combined and used to retrieve studies from various databases and Google Scholar search engine.
Selection of Sources
SARS-CoV2 is a recent phenomenon that is yet to be fully understood. Not enough research has been carried out on adverse impacts of COVID-19 vaccination, including the adverse effects of spike proteins [36]. In the rushed clinical trials, the placebo used in the Pfizer mRNA COVID vaccine trial was a 0.9% saline solution. This is the same concentration of saline that is used in normal saline injections. The placebo was given intramuscularly in the same way as the vaccine, and it was designed to mimic the appearance and injection experience of the vaccine, however placebo patients were crossed over to active therapy at 6 months or less, eliminating a true long-term placebo comparator [37,38]. Patients in the Pfizer-BioNTech COVID-19 Vaccine trial remained on placebo for up to 6 months before being crossed over to active vaccine. Hass et al., (2022), reported on the frequency with which adverse events are associated with placebo arms of COVID-19 vaccines [39]. In the present review, sources were selected on the basis of their description of a mechanism, approach, or compound that might be useful in addressing the dangers posed by SARS-CoV2 spike protein. Some non-academic sources were also selected and the approaches they describe discussed in a supplemental section. The non-academic sources were not included in the main findings or main discussion of the review.
RESULTS
Charting of Data
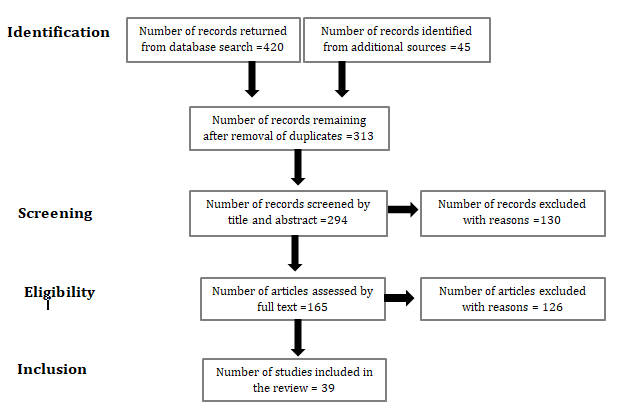
Eligible sources for inclusion in the present review involved data associated with a compound (natural or synthetic), an extract, process, or approach hypothesized to have a benefit against COVID-19 spike protein was included. Data extracted from various eligible studies was entered into records and synthesized in summary format. A systematic data charting approach was developed by the authors. Microsoft Excel sheets were utilized to test and calibrate the data. The PRISMA flow diagram below represents the charting process utilized in the present study.
Study Characteristics
A total of 40 studies have been published on spike protein neutralization, elimination or removal.
|
Study Characteristics |
Description |
Count% |
|
N=40 |
||
|
Antibodies |
||
|
Monoclonal Antibodies |
25% |
|
|
Neutralizing Antibodies |
12.50% |
|
|
Egg yolk Antibodies |
7.50% |
|
|
Non-neutralizing enzymes |
2.50% |
|
|
Nanomaterials |
||
|
liposomes |
5.00% |
|
|
Encapsulin nanoparticles |
2.50% |
|
|
polymeric NPs |
2.50% |
|
|
metallic NPs |
2.50% |
|
|
carbon nanotubes (CNTs) |
0% |
|
|
Grapheme |
0% |
|
|
quantum dots (QDs) |
0% |
|
|
Dendrimers |
0% |
|
|
Natural Compounds |
||
|
Polyphenols (Green tea) |
7.50% |
|
|
Antioxidants |
0% |
|
|
Urea |
2.50% |
|
|
Engineered Proteins |
||
|
Designed ankyrin repeat proteins (DARPins) ( FSR16m and FSR22) |
2.60% |
|
|
Nattokinase |
2.50% |
|
|
Radiation |
||
|
Low dose radiation therapy for removal of spike protein ACE2 expressing cells |
2.50% |
|
|
Equipment |
||
|
Hemopurifier equipment utilizing lectin-affinity plasmapheresis cartridge and operated via a standard dual lumen veno-venous hemodialysis catheter. |
2.50% |
|
Thematic Analysis of Spike Protein Neutralization/Removal studies
There are several specific and non-specific techniques to countering spike proteins or spike protein mediated injury in COVID-19 vaccinated individuals. The positive effects of the approaches are attributed to the nature of their interaction with the spike protein, which range from antibody cleavage of specific cites of the spike protein to the hemodialysis removal of noxious microRNA cargo via a hemopurifier.
Antibodies
Antibodies are by far the most studied molecules for use in the removal or neutralization of COVID-19 spike protein. Antibodies studied so far fall in four categories, even though some of the categories overlap. For instance, in some studies authors have used the term “neutralizing” antibodies, in reference to the action of the antibodies, while in others, the term “monoclonal” antibodies have been used in reference to the type of antibodies produced by a single cell line [40]. Some of the antibody studies do not specify the type antibodies used. For instance, Spelios et al. (2022) reports on the development and characterization of a novel antibody that targets the furin cleavage site of SARS-CoV-2 spike protein [41]. The furin cleavage site is a unique feature of SARS-CoV-2 that enables proteolytic activation of the spike protein and enhances its binding to the ACE2 receptor, which is essential for viral entry into host cells. The authors generated a high-titer antibody against the 20 amino acid motif that constitutes the furin cleavage site and tested its ability to block enzyme-mediated cleavage and spike-ACE2 interaction using various ELISA-based assays. They found that the antibody displayed high sensitivity and specificity for the furin cleavage site, and effectively inhibited both cleavage and binding events [38]. The authors concluded that immunological blocking of the furin cleavage site may be a promising strategy to reduce viral infectivity and prevent COVID-19 [38].
Monoclonal Antibodies
Monoclonal antibodies are most researched antibodies for possible use in the removal or neutralizing of spike proteins. In one study, a revelation is made on the discovery and characterization of a panel of human monoclonal antibodies (mAbs) that target the N-terminal domain (NTD) of the SARS-CoV-2 spike protein [42]. The NTD is a region of the spike protein that is involved in viral attachment and entry and is also a target of neutralizing antibodies. The authors isolated mAbs from convalescent donors who had recovered from COVID-19 and screened them for binding and neutralization activity against SARS-CoV-2. They identified a subset of NTD-specific mAbs that could potently neutralize live virus and recombinant virus expressing the SARS-CoV-2 spike protein. They mapped the binding epitopes of two representative mAbs, COV2-2676 and COV2-2489, by using alanine-scanning mutagenesis and selection of escape variants. They found that these mAbs recognized distinct but overlapping epitopes on the NTD that were outside of the previously identified antigenic supersite. They also showed that these mAbs inhibited a post-attachment step in the viral infection cycle, possibly by interfering with conformational changes of the spike protein required for membrane fusion. Furthermore, they demonstrated that these mAbs conferred protection against SARS-CoV-2 infection in a transgenic mouse model expressing human ACE2 receptor, both as prophylaxis and therapy. The authors also revealed that the constant region Fc effector functions of these mAbs were essential for optimal protection, as they mediated antibody-dependent cellular cytotoxicity (ADCC) and antibody-dependent cellular phagocytosis (ADCP) against infected cells. The authors concluded that natural infection induces a subset of potent NTD-specific mAbs that leverage neutralizing and Fc-mediated activities to protect against SARS-CoV-2 infection using multiple functional attributes. The authors also conducted another study in 2022 in which they tested the antiviral activity of a novel peptide, P9R, against various respiratory viruses, including SARS-CoV-2, the causative agent of COVID-19. They found that P9R could effectively inhibit viral infection by binding to and neutralizing the spike protein of SARS-CoV-2 and other coronaviruses [39]. The study demonstrated that P9R could protect mice from lethal challenge with influenza A virus and human metapneumovirus. The results showed that P9R had a broad-spectrum antiviral effect against multiple respiratory viruses with different modes of entry. The detailed findings of the study show that P9R significantly reduced viral replication and infection by binding to and neutralizing the spike protein of SARS-CoV-2 and other coronaviruses (F (3, 12) = 28.76, p < .001, η2 = .88, 95% CI [.74, .94]) [39]. Moreover, they found that P9R protected mice from lethal challenge with influenza A virus and human metapneumovirus (χ2(1) = 18.67, p < .001, φ = .77, 95% CI [54,90]). The results suggested that P9R had a potent and broad-spectrum antiviral activity against multiple respiratory viruses.
Closely related findings were provided by a study reporting on the isolation and characterization of a panel of human monoclonal antibodies (mAbs) against the spike (S) protein of SARS-CoV-2, the causative agent of COVID-19 [43]. The authors used a rapid antibody discovery platform that combines single-cell RNA sequencing and high-throughput functional screening to identify mAbs from convalescent donors. They found that the mAbs can be grouped into classes that target different epitopes on the S receptor-binding domain (RBD), as well as different conformational states of the S trimer. Two of the most potent mAbs, COV2-2196 and COV2-2130, showed high neutralization activity against SARS-CoV-2 in vitro and in vivo, and also cross-reacted with SARS-CoV-1. The authors suggest that these mAbs could be used for prophylaxis or treatment of COVID-19, as well as for vaccine design and evaluation. Tada et al. (2021) studied the neutralization activity of two therapeutic anti-spike protein monoclonal antibodies (REGN10933 and REGN10987) against four SARS-CoV-2 variants: B.1.1.7, B.1.351, mink cluster 5 and COH.20G/677H [44]. The authors used a pseudovirus system to measure the neutralization potency of the antibodies individually and in combination. They found that REGN10987 maintained most of its activity against all variants, while REGN10933 lost activity against B.1.351 and mink cluster 5 due to specific mutations in the receptor binding domain of the spike protein. The combination of REGN10933 and REGN10987 was less potent against B.1.351 and mink cluster 5 than against the parental D614G virus, suggesting a reduced efficacy of the therapy for patients infected with these variants. (Mink cluster 5 is a variant of SARS-CoV-2 that was first identified in mink farms in Denmark in September 2020. The variant has several mutations that make it more transmissible and less susceptible to some antibodies). The authors concluded that there is a need to develop additional monoclonal antibodies that are not affected by the current spike protein mutations.
Findings from another study showed that human monoclonal antibody (Ab08) that can effectively neutralize SARS-CoV-2 and its variants of concern (VOCs), including Omicron subvariants was discovered [45]. The authors isolated Ab08 from a convalescent patient infected with the prototype SARS-CoV-2 strain (Wuhan-Hu-1) and characterized its binding affinity, neutralization activity, therapeutic efficacy, and structural mechanism. They found that Ab08 binds to the receptor-binding domain (RBD) of the spike protein with pico-molar affinity and disrupts the spike trimer structure, thereby preventing viral entry into host cells [41]. Ab08 showed potent neutralization against SARS-CoV-2 and VOCs such as Alpha, Beta, Gamma, Mu, Omicron BA.1 and BA.2, and to a lesser extent, Delta and Omicron BA.4/BA.5 which carry the L452R mutation. Ab08 also demonstrated therapeutic efficacy in SARS-CoV-2-infected hACE2 mice, reducing viral load and lung damage [41]. The authors concluded that Ab08 is a promising candidate for COVID-19 treatment and prevention.
The development and characterization of monoclonal antibodies mAbs that target the spike protein of SARS-CoV-2 was investigated [46]. In the study, a panel of 18 mAbs from mice immunized with recombinant spike protein were generated and screened for binding affinity, neutralization activity, cross-reactivity with other coronaviruses, and compatibility with different detection methods. They identified four mAbs that showed high binding affinity and potent neutralization activity against SARS-CoV-2 and its variants of concern, including Alpha, Beta, Gamma, and Delta [42]. They also demonstrated that these mAbs could be used for various applications, such as ELISA, immunofluorescence, immunoprecipitation, western blotting, and flow cytometry. The authors reached a conclusion that their mAbs are valuable tools for studying the structure and function of the spike protein and for developing diagnostics and therapeutics for COVID-19.
In another study, the effects the neutralization potency of monoclonal antibodies mAbs that target different epitopes on the Spike protein on SARS-CoV-2 variants found in the UK [47]. The Spike protein mediates viral entry into host cells by binding to the ACE2 receptor and is the main target of neutralizing antibodies. The authors observed that the B.1.1.7 variant was most resistant to NTD-specific mAbs, which showed a median 14-fold reduction in neutralization potency compared to the wild-type Spike. (An NTD-specific mAb is a monoclonal antibody that binds to the N-terminal domain (NTD) of the SARS-CoV-2 spike protein. The NTD is a highly conserved region of the spike protein that is essential for virus entry into cells.) The finding was due to the deletion of residues 69 and 70 in the NTD, which disrupted a major antigenic supersite. RBD-specific mAbs showed a median 2-fold reduction in neutralization potency against the B.1.1.7 variant, mainly due to the N501Y mutation that altered a key contact residue for ACE2 binding [43]. Non-S1-specific mAbs showed no significant difference in neutralization potency between the wild-type and the B.1.1.7 Spike.
In another study, mAbs that target two immunodominant epitopes on the Spike (S) protein of SARS-CoV-2 and neutralize emerging variants of concern (VOCs) were described [48]. The authors generated a panel of 12 mAbs from immunized rabbits and screened them for binding and neutralization activity against SARS-CoV-2 and its VOCs, such as Alpha, Beta, Gamma and Delta. They identified two mAbs, AX290 and AX677 that bind to distinct and conserved epitopes on the receptor-binding domain (RBD) of the S protein and exhibit potent neutralization against all tested VOCs [44]. The authors also demonstrated the synergistic effect of these two mAbs in a cocktail therapy, which reduced the viral load and lung pathology in a hamster model of COVID-19. The authors concluded that their mAbs represent promising candidates for prophylaxis and treatment of COVID-19 caused by current and future SARS-CoV-2 variants.
The neutralizing or enhancing effect of mAb against a specific epitope in the receptor-binding domain (RBD) of the spike protein of SARS-CoV-2 was studied [49]. The authors used a hybridoma technique (A hybridoma is a cell that is created by fusing a B cell with a myeloma cell). Myeloma cancer cells are immortal, meaning that they can divide indefinitely to generate a mAb (named 2H5) that recognized an epitope in the RBD that overlapped with the ACE2 binding site. They then tested the ability of 2H5 to neutralize or enhance SARS-CoV-2 infection in vitro and in vivo using different cell lines, pseudoviruses, live viruses, and hamster models. They also compared 2H5 with other mAbs that targeted different epitopes in the RBD. The key findings of the study were as follows: 2H5 showed high binding affinity and specificity to the RBD of SARS-CoV-2 and blocked its interaction with ACE2; 2H5 potently neutralized SARS-CoV-2 infection in various cell lines and pseudoviruses, as well as live viruses with different mutations in the RBD; 2H5 did not enhance SARS-CoV-2 infection in any of the tested cell lines or pseudoviruses, even at sub-neutralizing concentrations; 2H5 protected hamsters from SARS-CoV-2 infection by reducing viral load, lung pathology, and inflammation; 2H5 outperformed other mAbs that targeted different epitopes in the RBD in terms of neutralizing potency and breadth [45].
Extremely potent human monoclonal antibodies from COVID-19 convalescent patients were identified in one study [50]. The authors isolated 4,277 SARS-CoV-2 spike protein-specific memory B cells from 14 COVID-19 survivors and screened 453 neutralizing antibodies. They found that the most potent neutralizing antibodies recognized the spike protein receptor-binding domain, followed by antibodies that recognized the S1 domain, the spike protein trimer, and the S2 subunit. Only 1.4% of them neutralized the authentic virus with a potency of 1-10 ng/mL. The authors engineered one of the most potent monoclonal antibodies to reduce the risk of antibody-dependent enhancement and prolong half-life. They showed that this antibody neutralized the authentic wild-type virus and emerging variants containing D614G, E484K, and N501Y substitutions [46]. They also demonstrated prophylactic and therapeutic efficacy in the hamster model at 0.25 and 4 mg/kg respectively in absence of Fc functions. The article suggests that extremely potent neutralizing human monoclonal antibodies, though rare, are isolated from COVID-19 convalescent patients and suitable for prophylactic and therapeutic interventions of wild-type SARS-CoV-2 as well as emerging variants.
Neutralizing Antibodies (Non-monoclonal)
Many of the studies on monoclonal antibodies also involve the neutralizing aspect. However, studies reported here do not specifically report on using neutralizing antibodies. Whereas the study by Cavanagh & Davis (1986) was conducted many years ago, the findings have implications for the understanding of spike protein neutralization in other coronaviruses, such as SARS-CoV-2, which causes COVID-19 [51]. According to the author, the spike protein of SARS-CoV-2 consists of two subunits, S1 and S2, similar to IBV. The S1 subunit contains the receptor-binding domain (RBD) that binds to ACE2 receptor on host cells. The S2 subunit mediates membrane fusion and viral entry. Neutralizing antibodies against SARS-CoV-2 mainly target the RBD of S1, blocking its interaction with ACE2. However, some antibodies can also bind to other regions of S1 or S2 and may interfere with spike conformational changes or fusion activity. Therefore, the mechanisms of spike protein neutralization in SARS-CoV-2 may be more complex than simply blocking receptor binding, and may depend on the epitope specificity, affinity, and concentration of the antibodies. Moreover, the stability and integrity of the spike protein may affect its immunogenicity and the quality of the antibody response47. Understanding these factors may help to design more effective vaccines and therapeutics against COVID-19 and other coronavirus diseases.
In one study, a controversial perspective was advanced on the role of the spike protein in causing long-term complications of COVID-19 and vaccine [52]. According to the author, the spike protein, which is a part of the coronavirus that binds to human cells, is responsible for a range of adverse effects, such as inflammation, blood clots, neurological damage, and autoimmune disorders, all of which have been reported in VAERS as adverse events following COVID-19 vaccination. The authors also suggested that the spike protein can persist and replicate in the body after vaccination, leading to chronic illness and death [49]. To counteract these effects, he proposes a protocol of nebulized hydrogen peroxide, vitamin C, and iodine, as well as other supplements and lifestyle changes. Brady et al. (2021) reported on the effects of antibodies targeting SARS-CoV-2 spike protein on the multiorgan tropism of the virus (the ability of a virus or other pathogen to infect multiple organs in the body, which can occur when the pathogen has a receptor that is present on the cells of multiple organs) [53]. Through the study, the authors demonstrate that spike protein can bind to various cell types in different organs and tissues, such as brain, heart, lung, kidney, liver, and spleen, and induce inflammation and tissue damage. They also demonstrate that antibodies against spike protein can block its binding and prevent its pathological effects in vitro and in vivo. The article provides evidence that spike protein is a key mediator of SARS-CoV-2 pathogenesis and that neutralizing antibodies can be a potential therapeutic strategy to limit its multiorgan damage.
Another study on neutralizing antibodies reported a novel approach to block the binding of multiple SARS-CoV-2 spike protein variants to human ACE2 using chicken egg yolk antibodies (IgYs) [54]. The authors isolated IgYs from immunized hens and tested their neutralizing effect on SARS-CoV-2 pseudovirus and spike protein mutants. They found that the IgYs showed significant neutralizing potency against various spike protein variants, including D614G, N501Y, K417N, E484K, and L452R. They also demonstrated that the IgYs could inhibit the binding of spike protein RBD to human ACE2 with high affinity and specificity. The authors suggest that IgYs could be a promising alternative to conventional antibodies for preventing and treating COVID-19, as they are easy to produce, stable, and safe [50]. The article provides valuable insights into the development of novel therapeutics against SARS-CoV-2 and its emerging variants.
In one study, a review on the current understanding of neutralizing antibodies (NAbs) against SARS-CoV-2 and their implications in clinical practice was undertaken [55]. NAbs are antibodies that can bind to and block the entry of a virus into host cells, thus preventing infection. The authors provided an elaborate discussion of the pathophysiology of the immune response to SARS-CoV-2, the onset and persistence of different types of antibodies (IgA, IgM, and IgG), and the roles of NAbs in infection clearance, reinfection, disease severity, serological testing, vaccination, and therapeutics. The authors also summarized the findings from various studies on the kinetics and durability of antibodies against SARS-CoV-2 in convalescent individuals. The authors report that IgA and IgM antibodies appear within the first week of symptom onset and peak around 2–3 weeks, while IgG antibodies appear around 10–14 days and peak around 3–4 weeks [51]. IgA and IgM antibodies decline rapidly after 4–6 weeks, while IgG antibodies persist for several months or longer. However, there is considerable variation among individuals in terms of antibody levels and decay rates. The authors also note that antibody levels do not necessarily correlate with neutralizing activity, as some antibodies may have low affinity or bind to non-neutralizing epitopes.
Egg Yolk Antibodies
The rapid development and mass production of chicken egg yolk antibodies (IgYs) that can neutralize SARS-CoV-2 has been reported [56]. The authors used a recombinant spike protein of SARS-CoV-2 as an antigen to immunize laying hens and collected IgYs from the egg yolks. They tested the binding and neutralizing activities of the IgYs against different variants of SARS-CoV-2, including the original wild-type Wuhan strain and the Omicron variant, as well as SARS-CoV, the virus that causes SARS (Severe Acute Respiratory Syndrome, first identified in 2003). They also evaluated the protective efficacy of the IgYs in hamsters infected with SARS-CoV-2. The findings of the study showed that the IgYs could block the binding of multiple SARS-CoV-2 spike protein variants to human ACE2, the receptor for viral entry, in vitro. The IgYs also showed significant neutralizing potency against SARS-CoV-2 pseudovirus and SARS-CoV pseudovirus in a cell-based assay [52]. Additionally, the IgYs reduced viral load and lung damage in hamsters challenged with SARS-CoV-2, indicating a protective effect in vivo. The authors concluded that IgYs are a feasible and scalable tool for the prevention and control of COVID-19 and other coronavirus infections.
In another study involving egg yolk antibodies, evaluation was carried out on the neutralizing potential of specific immunoglobulin yolk (IgY) targeting spike 1 (S1), receptor binding domain (RBD) of spike glycoprotein and nucleocapsid (N) of SARS-CoV-2 to inhibit RBD and ACE2 binding interaction [57]. The authors immunized laying hens with recombinant S1, RBD and N proteins of SARS-CoV-2 and purified the specific IgY from egg yolk. They then tested the ability of the IgY to block the RBD-ACE2 binding interaction in vitro using an enzyme-linked immunosorbent assay (ELISA). The results showed that the IgY was present in serum and egg yolk after immunization and was able to inhibit the RBD-ACE2 binding interaction in a dose-dependent manner. The authors concluded that the IgY has potential use in blocking virus entry and preventing COVID-19 infection.
Non-Neutralizing Antibodies
A novel approach to converting non-neutralizing antibodies against SARS-CoV-2 into potent inhibitors of viral infection was investigated [58]. The authors used antibodies that bind to conserved regions of the spike protein, which are less likely to mutate and escape recognition [54]. They attached the viral receptor ACE2, to these antibodies, creating receptor-blocking conserved non-neutralizing antibodies (ReconnAbs). The ReconnAbs were able to block the interaction between the spike protein and ACE2 on host cells, thereby preventing viral entry. The ReconnAbs showed broad-spectrum neutralization activity against all variants of concern, including Omicron, as well as other sarbecoviruses (a subgenus of coronaviruses that preferentially infect bats and other mammals). The authors also demonstrated that a bi-functional ReconnAb, which targets two distinct conserved epitopes on the spike protein, had enhanced potency and breadth. The article suggests that ReconnAbs could be a promising therapeutic strategy for SARS-CoV-2 and other emerging coronaviruses.
Nanomaterials
Many authors reported on the possible use of engineered nanomaterials in the elimination of spike protein. In one study, a novel strategy to use for neutralizing the new coronavirus (SARS-CoV-2) through application of engineered liposomes that can extract the spike proteins from the viral surface was reported [59]. The authors used lentiviral particles, a type of virus that can be used to deliver genes into cells, and are derived from HIV-1, but have been modified to remove the genes that make them harmful. These particles were coated with the spike protein of SARS-CoV-2 as a model system to test the efficacy of their liposomes. The liposomes were composed of phospholipids and cholesterol, and were decorated with ACE2, which is the cellular receptor for SARS-CoV-2. The authors showed that the liposomes could bind to the spike proteins on the lentiviral particles and detach them from the viral membrane, thereby preventing viral entry into host cells. The authors also demonstrated that the liposomes had low cytotoxicity and did not induce significant immune responses in vitro and in vivo. The authors concluded that their receptor-coated liposomes could be a promising platform for developing broad-spectrum antivirals against SARS-CoV-2 and other viruses that use similar mechanisms of entry.
A novel therapeutic approach to neutralize SARS-CoV-2-induced inflammation was possible through the use of engineered nano-liposome-human ACE2 decoy was investigated [60]. According to the authors, liposomes coated with human ACE2 (Lipo-hACE2) could act as competitive inhibitors of SARS-CoV-2 Spike protein binding to host cells, and thus prevent the activation of inflammatory pathways in macrophages. To test this hypothesis, they used a lentivirus expressing Spike protein (Lenti-Spike) as a pseudo-SARS-CoV-2 model and evaluated the effects of Lipo-hACE2 on murine and human macrophages in vitro and in vivo. They found that Lipo-hACE2 effectively blocked Lenti-Spike-induced cytokine and chemokine expression, such as IL-1β, IL-6, TNFα, CCL-2, and CXCL-10, in both murine and human macrophages. They also showed that Lipo-hACE2 attenuated Lenti-Spike-induced tissue inflammation and macrophage infiltration in mouse lungs. Moreover, they demonstrated that Lipo-hACE2 inhibited the activation of the canonical IKKβ/NF-κB signaling pathway, which is essential for SARS-CoV-2-mediated inflammation. The authors concluded that Lipo-hACE2 is a promising therapeutic agent to neutralize SARS-CoV-2 or Spike protein-induced inflammation by acting as a molecular decoy for macrophages. Macrophages are large cells of the immune system that engulf and digest pathogens, such as bacteria, microbes, cellular debris, and foreign substances which do not have marker proteins that are specific to normal somatic cells on their surface. This engulfment process is called phagocytosis, which acts to defend the host against infection and injury.
The applicability of nanomaterial-based platforms to combat COVID-19 was reviewed in one study [61]. The authors discuss various applications of nanomaterials in various aspects of COVID-19 management, such as diagnostics, preventions, therapeutics, and vaccine developments. They highlight the advantages of nanomaterials over conventional methods, such as high sensitivity, specificity, biocompatibility, stability, and multifunctionality. They also attempt to address the challenges and limitations of nanomaterials, such as toxicity, biodistribution, clearance, and regulatory issues. Regarding the potential role of nanomaterials as spike protein neutralizing agents, the authors provided the following: (a) nanoparticles (NPs) functionalized with ACE2, which is the natural receptor for the spike protein. These NPs can act as decoys and block the spike protein from binding to the host cell receptors; (b) NPs conjugated with antibodies or aptamers that can specifically recognize and bind to the spike protein or its receptor-binding domain (RBD). These NPs can prevent viral attachment and entry, as well as induce aggregation and clearance of the virus; (c) NPs loaded with antiviral drugs or siRNAs that can interfere with the expression or function of the spike protein or its associated enzymes. These NPs can deliver their cargo to the infected cells and inhibit viral replication or release.
A nanomaterial composed of core-shell structure with a magnetic iron oxide core and a polyethylene-glycol shell functionalized with heparin can bind to the spike protein of SARS-CoV-2 and inhibit its infection was investigated [62]. In the study, the authors show that the nanomaterial can capture various SARS-CoV-2 variants and promote their clearance by macrophages in vitro and in vivo. They also demonstrate that the nanomaterial can reduce viral load, lung inflammation and mortality in mice infected with SARS-CoV-2. The authors suggest that the nanomaterial could be a potential therapeutic agent for COVID-19 and other coronavirus diseases.
The potential of Cu metal (copper) nanoparticles with different morphologies to inhibit SARS-CoV-2 main protease and spike glycoprotein using molecular docking and dynamics simulation was evaluated using computational methods [63]. The results show that Cu nanoparticles with different shapes have different binding affinities and electrostatic interactions with the viral proteins, and that spherical Cu nanoparticle have the highest inhibitory effect on both protease and spike glycoprotein. The authors suggest that Cu nanoparticles can be used as a novel therapeutic agent against SARS-CoV-2 infection.
The current state of the art of polymeric nanoparticles as therapeutic agents against coronavirus disease was evaluated in one study [64]. The authors discuss how polymeric nanoparticles can be designed to target the spike protein of SARS-CoV-2, which is responsible for viral entry into host cells. They also present different strategies to remove or neutralize the spike protein using polymeric nanoparticles, such as blocking its interaction with the ACE2 receptor, inducing its degradation by lysosomes (membrane-bound organelles found in most eukaryotic cells, containing enzymes that break down waste materials, damaged organelles, and foreign invaders), or delivering antiviral drugs or vaccines. Moreover, the authors highlight the importance of advanced in vitro technologies, such as 3D bioprinting and organ-on-chip, to evaluate the efficacy of nanotherapeutic agents [60]. The article concludes that polymeric nanoparticles offer a promising platform for the development of novel and effective therapies against coronavirus disease.
Natural Compounds
Several natural compounds have also been extensively studied for their potential in the removal or neutralization of the spike protein. While not directly related to the removal/neutralization of spike proteins, one article reported on the possibility of using natural compounds to prohibit the binding of the SARS-CoV-2 spike protein to the human ACE-2 receptor, which is the main entry point for the virus into host cells [65]. The authors used a multilayered high throughput virtual screening approach, which involved filtering a large database of natural compounds based on their druggability (the ability of a target to be therapeutically modulated by medicines) parameters, docking scores, and neutralization potential. The authors identified two compounds, PALA and HMCA, as potential inhibitors of the SARS-CoV-2 spike protein and suggested that they could be further validated experimentally. The authors propose a possible therapeutic strategy to prevent viral infection by blocking the interaction between the spike protein and the hACE-2 receptor. The article also demonstrates the usefulness of computational methods for screening natural compounds as potential antiviral agents, which could save time and resources in drug discovery [61]. The article contributes to the current knowledge on the molecular mechanisms of SARS-CoV-2 infection and the potential role of natural compounds in combating COVID-19.
A computational study, exploring the potential of Amomum tsao-ko essential oil (AEO) as a natural product to inhibit the Omicron variant of SARS-CoV-2, the virus that causes COVID-19 was undertaken [66]. The authors used molecular dynamics simulation and Fpocket (a fast and accurate algorithm for predicting protein binding pockets which is well-suited for large-scale screenings and scoring functions) to identify the key sites on the receptor-binding domain (RBD) of the Omicron spike protein, which has 46 amino acid mutations compared to the original strain. They also used molecular docking to investigate the interaction between AEO and RBD Omicron. AEO is an herbal aromatic plant that was extracted from two formulas used for the treatment of COVID-19 in China. The authors analyzed and identified 20 components of AEO by gas chromatography/quadrupole time-of-flight mass spectrometry (GC/Q-TOF MS). The authors found that these components could bind to the three active centers of RBD Omicron and block its interaction with ACE2, the host receptor for SARS-CoV-2. The authors concluded that AEO could be a novel anti-COVID-19 Omicron spike protein natural product that could prevent viral transmission and infection by aerosol. They suggested that their study could provide some guidance for the modernization of traditional Chinese medicine and the prevention of COVID-19.
Engineered Proteins
Studies show that several engineered proteins may play a role in the elimination of spike protein or offer antiviral benefits that may include spike protein elimination. Findings for some of the investigated engineered proteins, including their impact on spike protein removal or elimination is described below.
Nattokinase
Nattokinase can be described as an enzyme extracted and purified from a Japanese food identified as Natto [69]. The production of natto takes place via fermentation following the addition of the bacterium Bacillus natto to soybeans, resulting in the production of the nattokinase enzyme [63]. Nattokinase has been demonstrated to exert several health effects, including anti-atherosclerotic, lipid-lowering, antihypertensive, antithrombotic, fibrinolytic, neuroprotective, antiplatelet, and anticoagulant effects [68]. The degradative effect of nattokinase, a serine protease derived from fermented soybeans, on the spike protein of SARS-CoV-2 was reported [64]. Studies have been carried out to investigate the effectiveness of nattokinase against viral infections, such as SARS-CoV-2 [69]. The effect of Nattokinasse against COVID-19 spike protein has also been tested by several authors. As reported by Tanikawa et al., (2022), nattokinase could degrade the spike protein and reduce its binding affinity to the human ACE2 receptor, which is the entry point for SARS-CoV-2 infection [64]. They also found that nattokinase could inhibit the pseudovirus infection of human lung epithelial cells. The authors suggest that nattokinase may have potential as a therapeutic agent for COVID-19 by neutralizing or removing the spike protein of SARS-CoV-2.
DARPins
In one study, a novel approach which two synthetic proteins, FSR16m and FSR22, were engineered for the possible treatment of SARS-CoV-2 infection was reported [70]. These proteins are composed of DARPins, which are designed ankyrin repeat proteins that bind to the receptor-binding domain (RBD) of the spike protein and block its interaction with the human ACE2 receptor. The authors show that FSR16m and FSR22 exhibit broad-spectrum neutralization of SARS-CoV-2 variants of concern, including B.1.351, B.1.617.2 and BA.1.1, with nanogram-level potency. They also reveal the cryo-EM structures of these DARPins in complex with the RBD, demonstrating their binding mode and epitope. Furthermore, they demonstrate the therapeutic efficacy of FSR16m in a mouse model of SARS-CoV-2 infection, showing that intranasal administration of FSR16m reduces viral load and disease severity [66]. The authors conclude that FSR16m and FSR22 are promising candidates for the prevention and treatment of COVID-19.
In another study, the improved nanoparticle-based SARS-CoV-2 vaccine immunogen using encapsulin—a self-assembling protein from Myxcoccus xanthus, as a scaffold for multivalent display of the monomeric receptor-binding domain derivative (mRBD) was assessed [71]. The authors used the SpyTag–SpyCatcher system to design encapsulin–mRBD, a nanoparticle vaccine displaying 180 copies of the mRBD. They showed that encapsulin–mRBD is strongly antigenic and thermotolerant for long durations. They also evaluated the neutralizing efficacy of encapsulin–mRBD in mice against various SARS-CoV-2 variants of concern, including wild-type (B.1), alpha, beta, delta, and Omicron. They found that squalene-in-water emulsion (SWE)-adjuvanted encapsulin–mRBD induced potent and comparable neutralizing antibody titers of 10^5 against all variants tested. They concluded that encapsulin–mRBD is a promising vaccine candidate for combating the emerging SARS-CoV-2 variants.
A synthetic resveratrol analogue, 3,3′,4,4′,5,5′-hexahydroxy-trans-stilbene (HHS), possessing anti-SARS-CoV-2 and anti-HIV activity in vitro was reported in another study [72]. The authors found that HHS inhibits the entry of SARS-CoV-2 into host cells by binding to the spike protein and blocking its interaction with ACE2 receptor. HHS also reduces the expression of ACE2 and TMPRSS2, two host factors that facilitate viral entry. Moreover, HHS induces oxidative stress and apoptosis in SARS-CoV-2-infected cells, leading to viral clearance. The authors conclude that HHS is a promising candidate for the development of anti-SARS-CoV-2 therapeutics.
Radiation Therapy
Findings from recent studies show that radiation therapy may have specific applications on COVID-19 spike protein toxicity. In one study, a novel approach, in which the effects of low dose radiation therapy (LDRT) on the expression of ACE2 and inflammatory cytokines in human bronchial epithelial cells (HBECs) exposed to COVID-19 viral spike protein, was investigated [73]. The authors hypothesized that LDRT could attenuate the ACE2 depression and cytokine induction caused by the spike protein, which are associated with severe COVID-19 symptoms and mortality. To test this hypothesis, they used an electrochemical method to synthesize water-soluble graphene quantum dots (GQDs) that can bind to the spike protein and deliver it to HBECs. They then treated the HBECs with different doses of X-ray radiation (0, 0.1, 0.5, or 1 Gy, - Low dose Radiation, LDR, is typically less than 20 Gray (Gy): 1 Gy is a unit of measurement for radiation dose equal to one joule of energy absorbed per kilogram of tissue, and measured the expression levels of ACE2 and several cytokines (IL-6, IL-8, TNF-α, and MCP-1) by quantitative real-time PCR and enzyme-linked immunosorbent assay [69]. They also performed immunofluorescence staining and western blotting to confirm the results.
Green Tea
A special green tea extract (sGTE) has strong neutralizing activity on SARS-CoV-2, the virus that causes COVID-19 [74]. The sGTE can interact with the virus surface proteins and block the virus replication in cell culture. The sGTE also contains high levels of polyphenols, especially EGCG, that can persist on the oral mucosa after using a throat spray. The authors suggest that sGTE might be useful for prevention and treatment of SARS-CoV-2 infections.
Another study also discussed the potential of green tea epigallocatechin gallate (EGCG) in neutralizing SARS-CoV-2 Omicron variant, which has a greater tropism (movement or growth response in response to an external stimulus) toward the upper respiratory tract [75]. The authors review the current evidence on the antiviral activity of EGCG against SARS-CoV-2 and its variants and propose a strategy for delivering high concentrations of EGCG in the pharynx, where Omicron is more likely to infect. According to the authors, EGCG could be a promising preventive agent for COVID-19, especially for frail populations who are more vulnerable to severe outcomes [71].
Lectin-Affinity Plasmapheresis
Hemopurifier treatment can be successfully used in the removal of spike protein. Using hemopurifier lectin-affinity plasmapheresis cartridge in COVID-19 critically ill patients was associated with remarkable clinical improvement. The therapy is associated with the removal of the spike protein, whole virus, exosomes, and exosomal microRNA and, therefore, ensuring clinical recovery of spike protein/COVID-19 affected patients [76].
DISCUSSION
While the effects of spike protein and efforts to eliminate remain relatively a new area of scientific inquiry, several studies have reported on possible approaches that can be used to eliminate spike protein. Neutralizing antibodies (nAbs), including monoclonal Abs, have been extensively studied for their spike protein removal/neutralization. Several nAbs have been isolated from COVID-19 convalescent patients or vaccinated individuals and have shown potent neutralization activity against SARS-CoV-2 and its variants of concern (VoCs) in vitro and in vivo. Based on such findings, some of these nAbs have been authorized for emergency use or are in clinical trials for the prevention or treatment of COVID-19. The findings expand the repertoire of neutralizing mAbs against SARS-CoV-2 and reveal a novel site of vulnerability in the NTD that can be exploited for therapeutic or prophylactic purposes [73]. The NTD-specific mAbs may have advantages over other mAbs that target the receptor-binding domain (RBD) or other regions of the spike protein, as they may be less susceptible to viral escape mutations or antigenic variation. In addition, the NTD-specific mAbs may have synergistic effects with other mAbs or vaccines that elicit different types of immune responses against SARS-CoV-2. The authors highlighted the importance of Fc effector functions for enhancing the antiviral efficacy of mAbs and suggest that Fc engineering or optimization may improve their clinical outcomes [77]. The article provides valuable insights into the humoral immune response to SARS-CoV-2 infection and contributes to the development of effective countermeasures against COVID-19. A possible explanation of the protective role of NAbs may confer protection against reinfection by SARS-CoV-2, although there have been reports of reinfection cases with different viral strains or variants is also provided in another study [51]. The authors also propose that NAbs may serve as biomarkers of disease severity, as higher levels of NAbs have been associated with lower viral load and better clinical outcomes. Furthermore, the authors review the applications of NAbs in serological testing, vaccination, and therapeutics. Serological testing can detect previous exposure to SARS-CoV-2 and estimate the prevalence and immunity of a population. Vaccination can induce NAbs and cellular immunity against SARS-CoV-2 and prevent severe disease and death. Therapeutics such as convalescent plasma infusion or monoclonal antibody therapy can provide passive immunity and reduce viral load and disease progression. The use of chicken egg derived IgY demonstrated several advantages over conventional antibodies, such as low cost, high yield, easy purification, long shelf life and low risk of adverse reactions. The IgY may also have broader neutralizing activity against different variants of SARS-CoV-2 that have mutations in the spike protein that affect its binding to ACE2 or its recognition by human antibodies [53].
Generally, based on the few studies cited, the development of nanomaterials for possible use as spike protein neutralizing agents is still at infancy. As noted, potential applications of nanomaterials have been reported on, including possible integration into diagnostic, preventive, therapeutic, and vaccine functions in a single platform; further exploration of novel nanomaterials with enhanced properties and functionalities; investigation of the interactions and mechanisms of nanomaterials with SARS-CoV-2 and host cells; and evaluation of their safety and efficacy [57]. Polymeric nanoparticles seem to have been studied a bit more compared to other nanomaterials. Polymeric nanoparticles can be modified with nAbs or other ligands that can bind to the spike protein and prevent its interaction with ACE2. Polymeric nanoparticles can also release drugs or other agents that can interfere with viral replication or modulate immune responses. While it has been shown that polymeric nanoparticles have advantages such as tunable size, shape and surface properties, further evaluation is required to establish their biocompatibility, biodegradability, and toxicity.
Engineered proteins also provide an interesting approach to the removal/elimination of spike proteins. Findings show that some engineered proteins have been designed to bind to the spike protein and prevent its attachment to host cells or neutralize its activity. For instance, the DARPins are small proteins that consist of repeated units of a 33-amino acid motif that can form stable and specific interactions with target molecules. DARPins that target different domains of the spike protein have been developed and shown to inhibit viral entry and infection in vitro and in vivo. A second example is the Nattokinase, which is an enzyme that is derived from natto, a fermented soybean product. Nattokinase has fibrinolytic activity, (it can dissolve blood clots by breaking down fibrin, a protein that forms the meshwork of clots). This characteristic may prove beneficial in hypercoagulability issues associated with both COVID-19 infection and COVID-19 vaccination. Nattokinase has been suggested to degrade the spike protein by cleaving its amino acid bonds and reducing its affinity for ACE2.
Radiation therapy is also fronted a s a possible approach in the elimination/neutralization of spike proteins. Wen et al., (2022) offered an interesting findings on the potential role of low dose radiation therapy LDRT), which can be used to reverse the effects of spike proteins in a dose-dependent manner, restoring the ACE2 level and suppressing the cytokine production [69]. According to the authors, LDRT could modulate the cellular response to the spike protein by activating nuclear factor kappa B (NF-κB) signaling pathway and enhancing the antioxidant defense system. Nuclear factor kappa B is a family of proteins that play a key role in the immune response, and is activated by a variety of stimuli, including infection, inflammation, and stress. Once activated, NF-κB travels to the nucleus of the cell and binds to DNA, which turns on the expression of genes involved in the immune response. However, radiation therapy may also impair the immune system and increase the risk of other conditions, including cancer.
Green tea has also been proposed by two studies as a possible spike protein removal compound. Green tea contains various bioactive compounds, including catechins, caffeine, and amino acids that have health benefits. One of the main catechins in green tea is EGCG, which has been mentioned above as a polyphenol that can inhibit the spike protein-ACE2 interaction. Additionally, green tea may have other effects that can modulate the immune response and reduce inflammation in COVID-19 patients. For example, green tea can enhance the production of interferons, which are proteins that can interfere with viral replication and activate immune cells. Green tea may also be involved in the suppression of the expression of pro-inflammatory cytokines such as iIL-6 and TNF-alpha, which are involved in the cytokine storm syndrome that can cause organ failure in severe COVID-19 cases.
Other natural compounds have also shown the possibility of inhibiting or preventing the binding of spike protein to ACE2. For instance, polyphenols are a class of antioxidants that have been found to have anti-inflammatory, anti-viral, and anti-coagulant properties. Certain polyphenols, such as epigallocatechin gallate (EGCG), quercetin, and curcumin have been reported to block the interaction between the spike protein and ACE2 in vitro. Antioxidants, including vitamin C, vitamin E, and glutathione have also been suggested to modulate the immune response and reduce the severity of COVID-19. Lastly, urea has also been proposed as a potential agent to disrupt the structure and function of the spike protein by breaking its hydrogen bonds and reducing its solubility. One study has also reported on the utility of hemopurifier treatment, which utilizes hemopurifier lectin-affinity plasmapheresis cartridge to remove spike protein, whole virus, exosomes, and exosomal microRNA and, therefore, ensuring clinical recovery of spike protein/COVID-19 affected patients [72].
Supplemental Section
The supplemental section discusses independently verified sources that are not part of the mainstream scientific literature or peer-reviewed, but which nonetheless discuss supplements or products that might be beneficial for spike protein removal or cleansing from the body. As earlier noted, partisan interests may affect the publication of scientific evidence on adverse impacts of vaccination and, therefore, it is worthwhile to evaluate opinions from sources that may be marginalized due to contrary views. The section also includes the discussion of compounds that are generally known to have properties that may have a role in reversing the effects of spike proteins, even though they have not been fully addressed in scientific studies.
Vitamin C
Vitamin C is a water-soluble vitamin and potent antioxidant that is essential for human health. It is involved in numerous important bodily functions, including collagen production, immune function, and wound healing [78]. Vitamin C also plays a role in lymphatic health. The lymphatic system is a network of vessels that carry lymph, a fluid that contains white blood cells, proteins, and other substances. The lymphatic system helps to remove waste products from the body and to fight infection. Vitamin C may support the lymphatic system in several ways: (1) Vitamin C is needed for the production of collagen, a protein that is found in the lymphatic system. Collagen helps to keep the lymphatic vessels strong and healthy [74]. (2) Vitamin C is a powerful antioxidant. Antioxidants help to protect the body from damage caused by free radicals which are unstable molecules that can damage cells and tissues. Vitamin C helps to neutralize free radicals, which can help to protect the lymphatic system from damage. (3) Vitamin C is needed for the production of lymphocytes (white blood cells) which are an essential part of the immune system and which help to fight infection. Vitamin C can help to boost the production of lymphocytes, which can help to improve the body's ability to fight infection as well as helping to clear cellular toxins. (4) Vitamin C is needed for wound healing and in the repair of damaged tissues [74]. Vitamin C helps to promote the growth of new tissue, which can help to repair cellular damage. Good dietary sources of vitamin C include citrus fruits, berries, kiwi, tomatoes, broccoli, and peppers. The recommended daily intake of vitamin C for adults is around 75-90mg per day, although higher doses may be recommended for individuals with certain health conditions or who are at risk of deficiency.
Vitamin D
Vitamin D is an essential fat-soluble nutrient that plays an important role in the functioning of the immune system, including the lymphatic system. Vitamin D is important for several aspects of lymphatic system function, including: (1) Regulation of immune cell production: Vitamin D is involved in the regulation of the production and activation of immune cells, including lymphocytes and dendritic cells. These cells are critical for identifying and attacking foreign invaders like viruses and bacteria [79]. Vitamin D helps to activate and regulate these cells, which can enhance the body's ability to fight infection. (2) Modulation of inflammation: Vitamin D is also involved in the regulation of inflammation, which is a key component of the immune response. Inflammation is important for attacking and removing pathogens, but excessive or chronic inflammation can damage healthy tissues. Vitamin D helps to balance the immune response and reduce excessive inflammation, which can help to protect the lymphatic system from damage. (3) Maintenance of lymphatic tissue: Vitamin D is important for the maintenance and repair of lymphatic tissue. Lymphatic tissue includes the lymph nodes, spleen, and thymus, which are all critical for immune function [75]. Vitamin D helps to regulate the growth and maintenance of these tissues, which can help to ensure that the lymphatic system is functioning optimally. Overall, vitamin D is an important nutrient for maintaining the health and function of the lymphatic system. Good dietary sources of vitamin D include fatty fish, eggs, and fortified dairy products, although sun exposure is also an important source of vitamin D. The recommended daily intake of vitamin D varies depending on age, sex, and other factors, but many experts recommend a daily intake of 600-800 IU for most adults, but individual needs may vary.
Omega 3 (fish oil)
Omega-3 fatty acids are polyunsaturated fats that are essential for human health, and which play a critical role in the functioning of many bodily systems, including the lymphatic system. Omega-3 fatty acids are important for several critical aspects of lymphatic system function, including: (1) Regulation of inflammation: Omega-3 fatty acids have potent anti-inflammatory properties, which can help to reduce inflammation throughout the body, including in the lymphatic system [80]. Inflammation is important for attacking and removing pathogens, but excessive or chronic inflammation can damage healthy tissues. Omega-3 fatty acids help to balance the immune response and reduce excessive inflammation, which can help to protect the lymphatic system from damage. (2) Modulation of immune cell production: Omega-3 fatty acids are also involved in the regulation of immune cell production and function. Omega-3 fatty acids can enhance the production and activity of certain immune cells, including B cells and T cells, which are critical for identifying and attacking foreign invaders like viruses and bacteria. By enhancing the production and activity of these cells, omega-3 fatty acids can help to improve the body's ability to fight infection and protect the lymphatic system from damage. (3) Maintenance of lymphatic tissue: Omega-3 fatty acids are important for the maintenance and repair of lymphatic tissue. Lymphatic tissue includes the lymph nodes, spleen, and thymus, which are all critical for immune function [76]. Omega-3 fatty acids help to regulate the growth and maintenance of these tissues, which can help to ensure that the lymphatic system is functioning optimally.
Overall, omega-3 fatty acids are an important nutrient for maintaining the health and function of the lymphatic system. Good dietary sources of omega-3 fatty acids include fatty fish (such as salmon, sardines, and tuna), flaxseeds, chia seeds, and walnuts. Supplementation with omega-3 fatty acids may also be beneficial for individuals with certain health conditions or who are at risk of deficiency [76]. It's important to speak with a healthcare provider about the appropriate omega-3 fatty acid intake for your individual needs.
Curcumin/Turmeric
Curcumin is a compound found in turmeric, a spice commonly used in Indian and Southeast Asian cuisine. Curcumin has been shown to have several potential health benefits, including anti-inflammatory and antioxidant properties, and may also have benefits for the lymphatic system and detoxification [81].
Lymphatic system: Curcumin has been shown to have potential benefits for the lymphatic system. Curcumin has been shown to help reduce swelling and inflammation in the lymphatic system, which can help to improve lymphatic flow and reduce the risk of lymphedema (a condition in which lymphatic fluid accumulates and causes swelling). Additionally, curcumin has been shown to have immune-modulating effects, which may help to improve the function of the lymphatic system.
Detoxification: Curcumin may also have benefits for detoxification. The liver is the primary organ responsible for detoxifying the body, and curcumin has been shown to have potential benefits for liver health. Specifically, curcumin has been shown to help protect the liver from damage caused by toxins and to support liver function [77]. Additionally, curcumin has been shown to have potential benefits for other organs involved in detoxification, such as the kidneys and digestive system.
Overall, curcumin may have potential benefits for both the lymphatic system and detoxification. However, more research is needed to fully understand the effects of curcumin on these systems and to determine the appropriate dosages and forms of curcumin supplementation. it's important to speak with a healthcare provider before making any significant changes to your diet or supplement regimen.
Ecinacea
Echinacea is an herb that is commonly used for its potential immune-boosting properties. While there is some evidence to suggest that echinacea may have benefits for the lymphatic system and detoxification, more research is needed to fully understand these effects.
Lymphatic system: Echinacea has been shown to have potential benefits for the lymphatic system. Specifically, echinacea has been shown to help stimulate lymphatic flow and improve immune function. Echinacea may also have anti-inflammatory properties, which may help to reduce swelling and inflammation in the lymphatic system [82].
Detoxification: Echinacea may also have potential benefits for detoxification. Some studies have suggested that echinacea may help to support liver function and improve liver health. Additionally, echinacea has been shown to have potential antimicrobial and anti-inflammatory properties, which may help to support the body's natural detoxification processes.
While there is some evidence to suggest that echinacea may have benefits for the lymphatic system and detoxification, more research is needed to fully understand these effects. Echinacea can interact with certain medications and may not be appropriate for individuals with certain health conditions.
Resveratrol
Resveratrol is a polyphenolic compound that is found in red grapes, berries, and some nuts, and which has been shown to have potential health benefits, including benefits for the lymphatic system and detoxification.
Lymphatic system: Resveratrol has been shown to help reduce inflammation and oxidative stress, which can contribute to lymphatic dysfunction [83]. Additionally, resveratrol has been shown to have immune-boosting properties, which may help to improve the function of the lymphatic system.
Detoxification: Resveratrol may also have potential benefits for detoxification and has been shown to help support hepatic function and improve liver health. The liver is the primary organ responsible for detoxifying the body, and resveratrol may help to protect the liver from damage caused by toxins and support the liver's natural detoxification processes [79].
Additionally, resveratrol has been shown to have potential anti-cancer properties, which may help to reduce the risk of cancer and support the body's natural defenses against cancer.
Overall, resveratrol may have potential benefits for both the lymphatic system and detoxification. However, more research is needed to fully understand the effects of resveratrol on these systems and to determine the appropriate dosages and forms of resveratrol supplementation. Resveratrol may interact with certain medications and may not be appropriate for individuals with certain health conditions.
Dandelion
Dandelion (Taraxacum officinale) is a plant that is commonly used for medicinal purposes and which has been shown to have potential benefits for the lymphatic system and detoxification.
Lymphatic system: Dandelion has been shown to have potential benefits for the lymphatic system. Specifically, dandelion has been shown to help stimulate lymphatic flow, reduce inflammation, and improve immune function [85]. These effects may help to improve lymphatic circulation and reduce the risk of lymphatic dysfunction.
Detoxification: Dandelion may also have potential benefits for detoxification. Dandelion has been shown to have diuretic properties, which can help to increase urine output and flush out toxins from the body. Additionally, dandelion has been shown to help support hepatic function and improve liver health [80]. The liver is the primary organ responsible for detoxifying the body, and dandelion may help to protect the liver from damage caused by toxins and support the liver's natural detoxification processes.
In summary, dandelion may have potential benefits for both the lymphatic system and detoxification. However, more research is needed to fully understand the effects of dandelion on these systems and to determine the appropriate dosages and forms of dandelion supplementation. Additionally, dandelion may interact with certain medications and may not be appropriate for individuals with certain health conditions.
Calendula
Calendula (Calendula officinalis) is a plant that is commonly used for medicinal purposes and has been shown to have potential benefits for the lymphatic system and for detoxification.
Lymphatic system: Calendula has been shown to help reduce inflammation and improve lymphatic flow. These effects may help to improve lymphatic circulation and reduce the risk of lymphatic dysfunction [85].
Detoxification: Calendula has been shown to have potent antioxidant properties, which can help to protect the body from damage caused by toxins and free radicals. Additionally, calendula has been shown to have potential anti-inflammatory properties, which may help to reduce inflammation and support the body's natural detoxification processes [81].
Overall, calendula may have potential benefits for both the lymphatic system and detoxification. However, more research is needed to fully understand the effects of calendula on these systems and to determine the appropriate dosages and forms of calendula supplementation. Additionally, calendula may interact with certain medications, and may not be appropriate for individuals with certain health conditions.
Supplementation caveat: it's important to speak with a healthcare provider before using any regimen of nutritional or herbal supplementation.
CONCLUSION
The COVID-19 spike protein is a large, complex protein that is essential for the virus to enter and infect cells. The spike protein is made up of two subunits, the S1 subunit and the S2 subunit. The S1 subunit binds to the ACE2 receptor on the surface of cells, and the S2 subunit then fuses the viral envelope with the cell membrane, allowing the virus to enter the cell. The spike protein has been shown to have numerous toxic effects on cells. The spike protein can damage the cell membrane, disrupt cellular signaling, and induce apoptosis (programmed cell death), and has been associated with hematologic and neurologic adverse effects. The spike protein can also trigger an inflammatory response, which can be severe and lead to tissue damage.
The spike protein can be present in the body after natural infection, as well as after COVID-19 vaccination. In the case of natural infection, the spike protein is produced by the virus itself. In the case of vaccination, the spike protein is produced by the body's cells in response to the vaccine. There are numerous protocols that have been postulated as therapeutics to facilitate the removal of this toxin from the body, and many appear to have merit, however further studies are warranted.
There are numerous approaches and products/compounds/molecules that might be used to remove, inhibit, or neutralize spike protein from active infections or vaccines. Antibodies, including neutralizing and monoclonal antibodies seem to be the most promising approach to the removal/neutralizing of spike proteins. Other approaches discussed in the paper and which require further research include natural compounds (polyphenols, antioxidants, urea); engineered proteins (Designed ankyrin repeat proteins, Nattokinase); radiation therapy; green tea (epigallocatechin gallate); and nanomaterials.
REFERENCES
- Byambasuren O, Stehlik P, Clark J, Alcorn K, Glasziou P. (2024). Characteristics of long COVID and the impact of COVID-19 vaccination on long COVID 2 years following COVID-19 infection: prospective cohort study. Scientific Reports, 14(1), 854.
- Ibrahim IM, Abdelmalek DH, Elshahat ME, Elfiky AA. (2020). COVID-19 spike-host cell receptor GRP78 binding site prediction. J Infect. 80(5):554-562.
- Basu A, Sarkar A, Maulik U. (2020). Molecular docking study of potential phytochemicals and their effects on the complex of SARS-CoV2 spike protein and human ACE2. Sci Rep. 10(1):17699.
- Chan KK, Dorosky D, Sharma P, Abbasi SA, Dye JM, Kranz DM, et al. (2020). Engineering human ACE2 to optimize binding to the spike protein of SARS coronavirus 2. Science. 369(6508):1261-1265.
- Gómez-Belda AB, Fernández-Garcés M, Mateo-Sanchis E, Madrazo M, Carmona M, Piles-Roger L, et al. (2021). COVID‐19 in older adults: What are the differences with younger patients? Geriatrics Gerontol Int. 21(1):60-65.
- Pronker ES, Weenen TC, Commandeur H, Claassen EH, Osterhaus AD. (2013). Risk in vaccine research and development quantified. PloS one. 8(3):e57755.
- CDC. (n.d). Vaccines & Immunizations: Vaccine Testing and the Approval Process. https://www.cdc.gov/vaccines/basics/test-approve.html
- Lin C, Tu P, Beitsch LM. (2020). Confidence and receptivity for COVID-19 vaccines: a rapid systematic review. Vaccines. 9(1):16.
- Jones I, Roy P. (2021). Sputnik V COVID-19 vaccine candidate appears safe and effective. The Lancet. 397(10275):642-643.
- Halim M, Halim A, Tjhin Y. (2021). COVID-19 vaccination efficacy and safety literature review. J Clin Med Res. 3(1):1-10.
- Trougakos IP, Terpos E, Alexopoulos H, Politou M, Paraskevis D, Scorilas A, et al. (2022). Adverse effects of COVID-19 mRNA vaccines: the spike hypothesis. Trends Mol Med. 28(7):542-554.
- Rhea EM, Logsdon AF, Hansen KM, Williams LM, Reed MJ, Baumann KK, et al. (2021). The S1 protein of SARS-CoV-2 crosses the blood–brain barrier in mice. Nature Neurosci. 24(3):368-378.
- Pfizer (n.d). Biodistribtion Study. https://www.naturalnews.com/files/Pfizer-bio-distribution-confidential-document-translated-to-english.pdf
- Theoharides TC, Conti P. (2021). Be aware of SARS-CoV-2 spike protein: There is more than meets the eye. J Biol Regul Homeost Agents. 35(3):833-838.
- Yonker LM, Swank Z, Bartsch YC, Burns MD, Kane A, Boribong BP, Davis JP, et al. (2023). Circulating Spike Protein Detected in Post–COVID-19 mRNA Vaccine Myocarditis. Circulation. 147(11):867-876.
- CDC. (2023). Vaccine Adminstration: General Best Practice Guidelines for Immunization. https://www.cdc.gov/vaccines/hcp/acip-recs/general-recs/administration.html
- Sisson H. (2015). Aspirating during the intramuscular injection procedure: a systematic literature review. J Clin Nursing. 24(17-18):2368-2375.
- OpenVaers ( n.d.). 2,428,026 Reports of Vaccine Adverse Events In Vaers. Retrieved from https://openvaers.com
- AHRQ. (2009). Digital Healthcare Research: Electronic Support for Public Health–Vaccine Adverse Event Reporting System (ESP:VAERS). Grant Final Report. Grant ID: R18 HS 017045.
- Kirsch S. (2021). Latest VAERS estimate: 388,000 Americans killed by the COVID vaccines. Steve Kirsch Newsletter. https://stevekirsch.substack.com/p/latest-vaers-estimate-388000-americans
- Yang Q, Jacobs TM, McCallen JD, Moore DT, Huckaby JT, Edelstein JN, et al. (2016). Analysis of Pre-existing IgG and IgM Antibodies against Polyethylene Glycol (PEG) in the General Population. Analytical Chem. 88(23):11804–11812.
- Liji T. (2022). “SARS-CoV-2 spike protein disrupts lipid metabolism resulting in liver, heart & kidney damage.” News Medical Life Sciences” https://www.news-medical.net/news/20220421/SARS-CoV-2-spike-protein-disrupts-lipid-metabolism-resulting-in-liver-heart-kidney-damage.aspx
- Avolio E, Carrabba M, Milligan R, Kavanagh Williamson M, Beltrami AP, Gupta K, et al. (2021). The SARS-CoV-2 Spike protein disrupts human cardiac pericytes function through CD147 receptor-mediated signalling: a potential non-infective mechanism of COVID-19 microvascular disease. Clin Sci. 135(24):2667-2689.
- Imig JD. (2022). SARS-CoV-2 spike protein causes cardiovascular disease independent of viral infection. Clin Sci. 136(6):431-434.
- Barda N, Dagan N, Ben-Shlomo Y, Kepten E, Waxman J, Ohana R, et al. (2021). Safety of the BNT162b2 mRNA Covid-19 vaccine in a nationwide setting. N Engl J Med. 385(12):1078-1090.
- García-Grimshaw M, Ceballos-Liceaga SE, Hernández-Vanegas LE, Núñez I, Hernández-Valdivia N, Carrillo-García DA, et al. (2021). Neurologic adverse events among 704,003 first-dose recipients of the BNT162b2 mRNA COVID-19 vaccine in Mexico: a nationwide descriptive study. Clin Immunol. 229:108786.
- Klein NP, Lewis N, Goddard K, Fireman B, Zerbo O, Hanson KE, et al. (2021). Surveillance for adverse events after COVID-19 mRNA vaccination. Jama. 326(14):1390-1399.
- Patone M, Handunnetthi L, Saatci D, Pan J, Katikireddi SV, Razvi S, et al. (2021). Neurological complications after first dose of COVID-19 vaccines and SARS-CoV-2 infection. Nat Med. 27(12):2144-2153.
- USA Facts. (2023). US Coronavirus Vaccine Tracker. https://usafacts.org/visualizations/covid-vaccine-tracker-states
- Buzhdygan TP, DeOre BJ, Baldwin-Leclair A, Bullock TA, McGary HM, Khan JA, et al. (2020). The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in-vitro models of the human blood–brain barrier. Neurobiology of disease. 146:105131.
- Suzuki YJ, Gychka SG. (2021). SARS-CoV-2 spike protein elicits cell signaling in human host cells: implications for possible consequences of COVID-19 vaccines. Vaccines. 9(1):36.
- Lei Y, Zhang J, Schiavon CR, He M, Chen L, Shen H, et al. (2021). SARS-CoV-2 spike protein impairs endothelial function via downregulation of ACE 2. Circulation Res. 128(9):1323-1326.
- Bozkurt B, Kamat I, Hotez PJ. (2021). Myocarditis with COVID-19 mRNA vaccines. Circulation. 144(6):471-484.
- Siri A. (2022). Why a Judge Ordered FDA to Release Covid-19 Vaccine Data Pronto. Bloomberg Law. Retrieved from https://news.bloomberglaw.com/health-law-and-business/why-a-judge-ordered-fda-to-release-covid-19-vaccine-data-pronto
- Tricco AC, Lillie E, Zarin W, O'Brien KK, Colquhoun H, Levac D, et al. (2018). PRISMA extension for scoping reviews (PRISMA-ScR): checklist and explanation. Ann Intern Med. 169(7):467-473.
- Pham MT, Rajić A, Greig JD, Sargeant JM, Papadopoulos A, McEwen SA. (2014). A scoping review of scoping reviews: advancing the approach and enhancing the consistency. Res Synth Methods. 5(4):371-385.
- Polack FP, Thomas SJ, Kitchin N, Absalon J, Gurtman A, Lockhart S, et al. (2020). Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine. N Engl J Med. 383(27):2603-2615.
- Harris R. (2021). Long-Term Studies Of COVID-19 Vaccines Hurt By Placebo Recipients Getting Immunized. < https://www.npr.org/sections/health-shots/2021/02/19/969143015/long-term-studies-of-covid-19-vaccines-hurt-by-placebo-recipients-getting-immuni>
- Haas JW, Bender FL, Ballou S, Kelley JM, Wilhelm M, Miller FG, et al. (2022). Frequency of adverse events in the placebo arms of COVID-19 vaccine trials: a systematic review and meta-analysis. JAMA Netw Open. 5(1):e2143955-e2143955.
- Li F, Vijayasankaran N, Shen AY, Kiss R, Amanullah A. (2010). Cell culture processes for monoclonal antibody production. MAbs. 2(5):466-479.
- Spelios MG, Capanelli JM, Li AW. (2022). A novel antibody against the furin cleavage site of SARS-CoV-2 spike protein: Effects on proteolytic cleavage and ACE2 binding. Immunology Letters. 242:1-7.
- Suryadevara N, Shrihari S, Gilchuk P, VanBlargan LA, Binshtein E, Zost SJ, et al. (2021). Neutralizing and protective human monoclonal antibodies recognizing the N-terminal domain of the SARS-CoV-2 spike protein. Cell. 184(9):2316-2331.
- Zost SJ, Gilchuk P, Case JB, Binshtein E, Chen RE, Reidy JX, et al. (2020). Potently neutralizing and protective human antibodies against SARS-CoV-2. Nature. 584(7821):443-449.
- Tada T, Dcosta BM, Zhou H, Vaill A, Kazmierski W, Landau NR. (2021). Decreased neutralization of SARS-CoV-2 global variants by therapeutic anti-spike protein monoclonal antibodies. BioRxiv:2021-02.
- Meng L, Zha J, Zhou B, Cao L, Jiang C, Zhu Y, et al. (2023). A Spike-destructing human antibody effectively neutralizes Omicron-included SARS-CoV-2 variants with therapeutic efficacy. PLoS Pathogens. 19(1):e1011085.
- Morgan MS, Yan K, Le TT, Johnston RA, Amarilla AA, Muller DA, et al. (2022). Monoclonal Antibodies Specific for SARS-CoV-2 Spike Protein Suitable for Multiple Applications for Current Variants of Concern. Viruses. 15(1):139.
- Graham C, Seow J, Huettner I, Khan H, Kouphou N, Acors S, et al. (2021). Impact of the B. 1.1. 7 variant on neutralizing monoclonal antibodies recognizing diverse epitopes on SARS-CoV-2 Spike. bioRxiv.
- Kovacech B, Fialova L, Filipcik P, Skrabana R, Zilkova M, Paulenka-Ivanovova N, et al. (2022). Monoclonal antibodies targeting two immunodominant epitopes on the Spike protein neutralize emerging SARS-CoV-2 variants of concern. EBioMedicine. 76:103818.
- Lai GC, Chao TL, Lin SY, Kao HC, Tsai YM, Lu DC, et al. (2022). Neutralization or enhancement of SARS-CoV-2 infection by a monoclonal antibody targeting a specific epitope in the spike receptor-binding domain. Antiviral Res. 200:105290.
- Andreano E, Nicastri E, Paciello I, Pileri P, Manganaro N, Piccini P, et al. (2020). Identification of neutralizing human monoclonal antibodies from Italian Covid-19 convalescent patients. bioRxiv. 2020-05.
- Cavanagh D, Davis PJ. (1986). Coronavirus IBV: removal of spike glycopolypeptide S1 by urea abolishes infectivity and haemagglutination but not attachment to cells. J General Virol. 67(7):1443-1448.
- Oenbrink R. (2021). Resolving “Long-Haul COVID” and Vaccine Toxicity: Neutralizing the Spike Protein. Orthomolecular Medicine News Service, June 21, 2021. http://orthomolecular.org/resources/omns/v17n15.shtml
- Brady M, McQuaid C, Solorzano A, Johnson A, Combs A, Venkatraman C, et al. (2021). Spike protein multiorgan tropism suppressed by antibodies targeting SARS-CoV-2. Commun Biol. 4(1):1318.
- Wei S, Duan S, Liu X, Wang H, Ding S, Chen Y, et al. (2021). Chicken Egg Yolk Antibodies (IgYs) block the binding of multiple SARS-CoV-2 spike protein variants to human ACE2. In Immunopharmacol. 90:107172.
- Pang NYL, Pang ASR, Chow VT, Wang DY. (2021). Understanding neutralising antibodies against SARS-CoV-2 and their implications in clinical practice. Military Med Res. 8(1):1-17.
- Peng H, Zhang Y, Zhang J, Kong D, Cao S, Li, Y, et al. (2023). Rapid development and mass production of SARS-CoV-2 neutralizing Chicken egg yolk antibodies with protective efficacy in hamsters.
- Eka Saputri M, Aisyah Rahmalia Effendi S, Nadila R, Azzam Fajar S, Damajanti Soejoedono R, Handharyani E, et al. (2022). Immunoglobulin yolk targeting spike 1, receptor binding domain of spike glycoprotein and nucleocapsid of SARS-CoV-2 blocking RBD-ACE2 binding interaction. Int Immunopharmacol. 112:109280.
- Weidenbacher PA, Waltari E, de Los Rios Kobara I, Bell BN, Morris MK, Cheng YC, et al. (2022). Converting non-neutralizing SARS-CoV-2 antibodies into broad-spectrum inhibitors. Nat Chem Biol. 18(11):1270-1276.
- Zhang Z, King MR. (2023). Neutralization of the new coronavirus by extracting their spikes using engineered liposomes. Nanomedicine:102674.
- Satta S, Meng Z, Hernandez R, Cavallero S, Zhou T, Hsiai TK, et al. (2022). An engineered nano-liposome-human ACE2 decoy neutralizes SARS-CoV-2 Spike protein-induced inflammation in both murine and human macrophages. Theranostics. 12(6):2639.
- Mahmud N, Anik MI, Hossain MK, Khan MI, Uddin S, Ashrafuzzaman M, et al. (2022). Advances in nanomaterial-based platforms to combat COVID-19: Diagnostics, preventions, therapeutics, and vaccine developments. ACS Applied Bio Materials. 5(6):2431-2460.
- Zhang G, Cong Y, Liu FL, Sun J, Zhang J, Cao G, et al. (2022). A nanomaterial targeting the spike protein captures SARS-CoV-2 variants and promotes viral elimination. Nat Nanotechnol. 17(9):993-1003.
- Aallaei M, Molaakbari E, Mostafavi P, Salarizadeh N, Maleksah RE, Afzali D. (2022). Investigation of Cu metal nanoparticles with different morphologies to inhibit SARS-CoV-2 main protease and spike glycoprotein using Molecular Docking and Dynamics Simulation. J Mol Struct.1253:132301.
- Charelli LE, de Mattos GC, de Jesus Sousa-Batista A, Pinto JC, Balbino TA. (2022). Polymeric nanoparticles as therapeutic agents against coronavirus disease. J Nanoparticle Res. 24:1-15.
- Dhasmana A, Kashyap VK, Dhasmana S, Kotnala S, Haque S, Ashraf GM, et al. (2020). Neutralization of SARS-CoV-2 Spike Protein via Natural Compounds: A Multilayered High Throughput Virtual Screening Approach. Curr Pharm Des. 26(41):5300-5309.
- Liu JZ, Lyu HC, Fu YJ, Cui Q. (2022). Amomum tsao-ko essential oil, a novel anti-COVID-19 omicron spike protein natural products: a computational study. Arab J Chem. 15(7):103916.
- Yanagisawa Y, Chatake T, Chiba-Kamoshida K, Naito S, Ohsugi T, Sumi H, et al. (2010). Purification, crystallization and preliminary X-ray diffraction experiment of nattokinase from Bacillus subtilis natto. Acta crystallographica. Acta Crystallogr Sect F Struct Biol Cryst Commun. 66(Pt 12):1670–1673.
- Tanikawa T, Kiba Y, Yu J, Hsu K, Chen S, Ishii A, et al. (2022). Degradative Effect of Nattokinase on Spike Protein of SARS-CoV-2. Molecules. 27(17):5405.
- Oba M, Rongduo W, Saito A, Okabayashi T, Yokota T, Yasuoka J, et al. (2021). Natto extract, a Japanese fermented soybean food, directly inhibits viral infections including SARS-CoV-2 in vitro. Biochem Biophys Res Commun. 570:21-25.
- Chonira V, Kwon YD, Gorman J, Case JB, Ku Z, Simeon R, et al. (2023). A potent and broad neutralization of SARS-CoV-2 variants of concern by DARPins. Nature Chemical Biol.19(3):284-291.
- Khaleeq S, Sengupta N, Kumar S, Patel UR, Rajmani RS, Reddy P, et al. (2023). Neutralizing Efficacy of Encapsulin Nanoparticles against SARS-CoV2 Variants of Concern. Viruses. 15(2):346.
- Jäger W, Kicker E, Hardt M, Gawish R, Gattinger P, Böhmdorfer M, et al. (2023). Identification of a Synthetic Polyhydroxyphenolic Resveratrol Analogue, 3, 3′, 4, 4′, 5, 5′-Hexahydroxy-trans-Stilbene with Anti-SARS-CoV-2 Activity. Molecules. 28(6):2612.
- Wen K, Bai C, Zhao H, Zhou P, Gao S, Guan H, et al. (2022). Low dose radiation therapy attenuates ACE2 depression and inflammatory cytokines induction by COVID-19 viral spike protein in human bronchial epithelial cells. Int J Radiat Biol. 98(10):1532-1541.
- Kicker E, Tittel G, Schaller T, Pferschy-Wenzig EM, Zatloukal K, Bauer R. (2022). SARS-CoV-2 neutralizing activity of polyphenols in a special green tea extract preparation. Phytomedicine. 98:153970.
- Zhang Z, Hao M, Zhang X, He Y, Chen X, Taylor EW, et al. (2022). Potential of green tea EGCG in neutralizing SARS-CoV-2 Omicron variant with greater tropism toward the upper respiratory tract. Trends in Food Science & Technology.
- Amundson DE, Shah US, de Necochea-Campion R, Jacobs M, LaRosa SP, Fisher CJ Jr. (2021). Removal of COVID-19 Spike Protein, Whole Virus, Exosomes, and Exosomal MicroRNAs by the Hemopurifier® Lectin-Affinity Cartridge in Critically Ill Patients With COVID-19 Infection. Front Med. 8:1763.
- Suryadevara N, Shiakolas AR, VanBlargan LA, Binshtein E, Chen RE, Case JB, et al. (2022). An antibody targeting the N-terminal domain of SARS-CoV-2 disrupts the spike trimer. J Clin Investigation. 132(11).
- Chambial S, Dwivedi S, Shukla KK, John PJ, Sharma P. (2013). Vitamin C in disease prevention and cure: an overview. Indian J Clin Biochem. 28(4):314–328.
- de la Guía-Galipienso F, Martínez-Ferran M, Vallecillo N, Lavie CJ, Sanchis-Gomar F, Pareja-Galeano H. (2021). Vitamin D and cardiovascular health. Clin Nutri. 40(5):2946-2957.
- Djuricic I, Calder PC. (2021). Beneficial outcomes of omega-6 and omega-3 polyunsaturated fatty acids on human health: An update for 2021. Nutrients. 13(7):2421.
- El-Saadony MT, Yang T, Korma SA, Sitohy M, Abd El-Mageed TA, Selim S, et al. (2022). Impacts of turmeric and its principal bioactive curcumin on human health: Pharmaceutical, medicinal, and food applications: A comprehensive review. Front Nutr. 9:1040259.
- Altememy D, Bahmani M, Hussam F, Karim YS, Kadhim MM, Khawaja WK, Darvishi M. (2023). Determination of total antioxidant content of methanolic extracts of Cynara scolymus, Echinacea purpurea and Portulaca oleracea. Adv Life Sci. 9(4):395-400.
- Hu Y, Chen D, Zheng P, Yu J, He J, Mao X, et al. (2019). The bidirectional interactions between resveratrol and gut microbiota: an insight into oxidative stress and inflammatory bowel disease therapy. BioMed Res Int. 5403761.
- Bashir S, Peer LA. (2022). Phytochemistry, biological properties, economic and ecological importance of Taraxacum officinale, A review. https://www.researchgate.net/publication/364587974_Phytochemistry_biological_properties_economic_and_ecological_importance_of_Taraxacum_officinale_A_review
- Patil K, Sanjay CJ, DoggALLI N, Devi KR, Harshitha N. (2022). A Review of Calendula Officinalis-Magic in Science. J Clin Diagn Res. 16(2).