Previous Issues Volume 2, Issue 1 - 2018
The Growth Cartilage and Beyond: Absence of Medullary Bone in Silver Carp Ribs
Soha A. Soliman*
Department of Histology, Faculty of Veterinary medicine, South Valley University, Qena, Egypt. Corresponding Author: Soha A. Soliman, Department of Histology, Faculty of Veterinary medicine, South Valley University, Qena, Egypt Tel: +47 67964599 E-Mail: [email protected]
Received Date:20 Sep 2018 Accepted Date: 2: 29 Sep 2018 Published Date: 03 Oct 2018
Copyright © 2018 Soliman AS
Citation: Soliman AS. (2018). The Growth Cartilage and Beyond: Absence of Medullary Bone in Silver Carp Ribs. Mathews J Cytol Histol. 2(1): 008.
ABSTRACT
The histological features of the ventral ribs were identified using semithin sections. The ventral rib had two cartilaginous ends and an elongated body. The cartilaginous ends exhibited typical zonal organization of the growth plate. The body was comprised of bone collar and the medullary cavity. Chondrocytes were organized in the resting, proliferating and hypertrophic zones. Cartilage matrix subjected to extensive degradation and erosion particularly by Chondroclasts. At the level of the hypertrophic zone, perichondral bone was formed and small projections of the bone tissues extended to the cartilage matrix. However, no medullary bone was formed beyond the hypertrophic zone. Cartilage erosion was also identified in the sub-osseous tissue and in the interior cartilage matrix. In the body of the ventral ribs, a remnant of the hypertrophic zone was surrounded by perichondral bone. Chondroclast was detected at the eroding surface of the cartilage matrix. Active osteoblasts, that had an oval or columnar in shape, were aligned at the surface of the bone collar as well as the resting osteoblasts. In conclusion, ventral ribs of the silver carp grow by perichondral bone formation in addition to the linear expansion of the growth cartilage.
INTRODUCTION
Skeletal system attracts biologists to investigate varieties of the morphological characteristics among different aquatic species. The significance of fish skeleton extends not only to support the body but also as aids for locomotion and swimming. Aquatic skeletal modification adopts diverse physical activities including the locomotion on land, swimming patterns, habitual behaviors such as migration. The supportive fish skeleton consists of different types of bone, cartilage and dentine and enamel/enameloid. Bone categorized according to the existence of cellular components. The cellular bone harbors typical organization of the osteocytic network. The acellular bone is devoid of osteocytes. Fish cartilage is classified depending on the predominant cellular or fibrillar components such as hyaline, Elastic/cell‐rich cartilage , Elastic hyaline cell cartilage , Fibro/cell‐rich cartilage , Fibrohyaline‐cell cartilage, Degrading cartilages and cartilage-like tissue including Chondroid bone, Mucochondroid and notochord [1].
Bone develops by two modes intramembranous ossification and endochondral ossification. Intramembranous ossification is characteristic for the cranial bone where the bone spicules are formed within the condensed mesenchymal membranes. Endochondral ossification depends on formation of a transient cartilage template which is gradually replaced by bone tissue. Cartilage template exhibits a specific zonal arrangement where the cartilage grows through progressive proliferation of chondrocytes. Proceeding the proliferating stage, the resting stage that retains the self-renewal capacity. Proliferating chondrocytes exit the cell cycle and undergo hypertrophy. Hypertrophied chondrocytes prepare the extracellular matrix for deposition of bone tissue. Endochondral bone formation archives bone growth as chondrocytes capable to replicate and produce extracellular matrix that adopt the bone formation.
Bone growth is widely investigated in mammals and avian species [2, 3, 4]. Few studies described bone growth in aquatic species. Some literatures described the two modes of bone growth in aquatic species [5, 6] as well as the perichondrial and parachondral type of ossification [1]. However, specification of the different modes of bone growth to the axial and appendicular skeleton has not yet been elucidated. Moreover, growth of different types of bone in different skeletal elements and species of fish is still ambiguous. The current study aimed to investigate type of ossification occurred in the ventral ribs of the silver carp.
MATERIAL AND METHODS
Fish transportation was performed according to [7]. Ten samples were obtained from silver carp, Hypophthalmichthys molitrix. The ventral ribs were carefully dissected from the fish body. Sampling and samples fixation were performed regarding to [3]. Protocol of processing for preparation of resin embedding samples was performed according to [4]. Semithin sections were stained using toluidine blue [7, 8].
RESULTS
The histological features of the ventral ribs were identified using semithin sections. The ventral rib had two cartilaginous ends and an elongated body. The cartilaginous ends exhibited typical zonal organization of the growth plate (Figure. 1A-C, E). The body was comprised of bone collar and the medullary cavity (Figure. 1D).
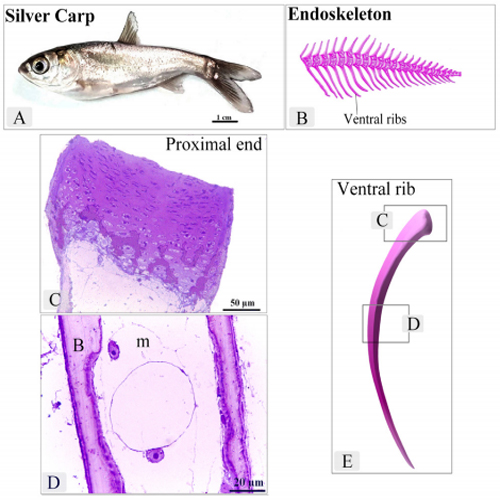
Figure 1: Location and General morphology of the ventral ribs in Silver carp
A : Silver carp. B: An illustration of a part of the endoskeleton of the Silver carp. C: proximal end of the ventral rib. Note the cartilage template exhibited typical zonal organization of the growth plate. D: body of the ventral rib. Note Bone collar (B), Medullary Cavity (m). E: An illustration of the ventral rib. note the squared areas of C and D represent the areas of Figure C and D.
Chondrocytes were organized in the resting, proliferating and hypertrophic zones. Cartilage matrix subjected to extensive degradation and erosion particularly by Chondroclasts (Figure. 2). At the level of the hypertrophic zone, perichondral bone was formed and small projections of the bone tissues extended to the cartilage matrix (Figure. 3). However, no medullary bone was formed beyond the hypertrophic zone. Cartilage erosion was also identified in the sub-osseous tissue and in the interior cartilage matrix (Figure. 4). In the body of the ventral ribs, a remnant of the hypertrophic zone was surrounded by perichondral bone. Chondroclast was detected at the eroding surface of the cartilage matrix. Active osteoblasts, that had an oval or columnar in shape, were aligned at the surface of the bone collar as well as the resting osteoblasts (Figure 5).
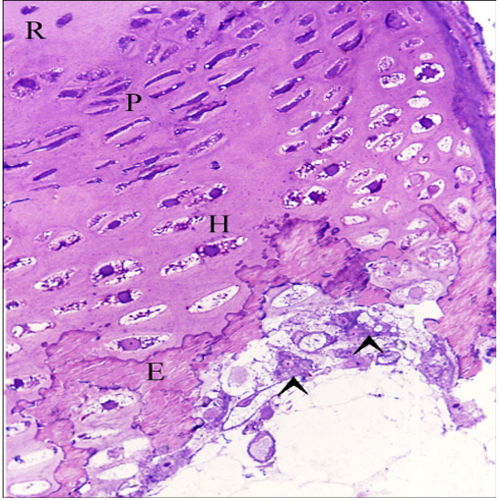
Figure 2: Cartilage template exhibited typical zonal organization of the growth plate Chondrocytes were organized in the resting (R), proliferating (P), Hypertrophic (H) zones. Note degradation and erosion (E) of the metachromatic matrix of growth cartilage. Chondroclast (arrowheads).
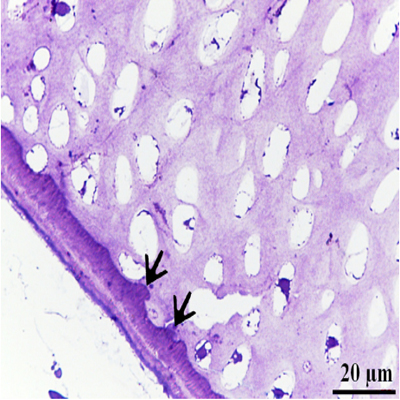
Figure 3: Perichondral bone occurred around the upper hypertrophic zone Hypertrophic chondrocytes (H), Note: Bone was confined to the perichondral area (B), small projections of the bone tissues into the cartilage matrix (arrows)
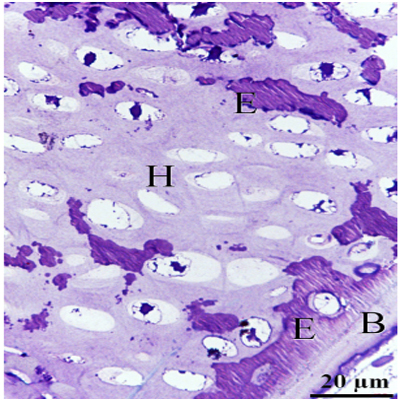
Figure 4: Erosion of the cartilage matrix in the lower hypertrophic zone Hypertrophic chondrocytes (H), Perichondral bone (B), Erosive (E) event was detected in the sub-osseous tissue and in the interior cartilage matrix.
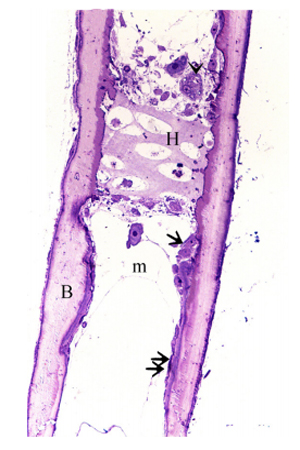
Figure 5: The body of the ventral rib Remnants of the hypertrophic zone (H), Perichondral bone (B), Chondroclast (arrowheads), active osteoblasts (arrow) had an oval or columnar shape, resting osteoblasts (double arrow) were flattened.
DISCUSSION
The functional significance of the ribs in fish render it worth to be studied. Fish ribs support the myosepta and forms the rib cage in which the vital organs are protected such as liver,spleen, heart [9, 10]. In the current study, one type of bone growth was identified in the ventral ribs of the silver carp. The ribs grow primarily through perichondral ossification. Perichondral ossification occurs in a manner identical to periosteal bone formation as described in mammals and avian species. The perichondrium undergoes maturation and transforms to periosteum in which the cells are driven toward the osteogenic cell lineage, activate genes characteristic of osteoblasts and secrete components of bone matrix around the circumference of the cartilage [11]. Perichondral ossification has been identified in medaka hypurals [12], gill cartilage of the Jordanella floridae [13, 14, 5].
Although typical growth cartilage was detected in the ends of the ventral ribs of the silver carp, it lacked endochondral ossification and medullary bone formation. Two modes of cartilage-dependent bone growth have been specified for fish; endochondral and perichondral ossification [1]. Endochondral bone develops by gradual replacement of the cartilage tissue by bone tissue and occurs via three mechanisms. The first mechanism is formation of the perichondral bone which is developed by an apposition manner. The second mechanism is formed via replacement of the cartilage through an endochondral ossification. The third mechanism is continuation of apposition bone formation [15]. Not only the mechanisms of cartilage replacement are different but also the type of bone is varied in different anatomical locations. Endochondral bone could be cellular or acellular types [5]
Occurrence of endochondral bone formation is probably related to fish size [1, 6]. Endochondral bone formation has been identified in large sized teleost species such as carp Cyprininus carpio, Atlantic salmon [16] but is uncommon for the smallsized species such as medaka [17] and Astatotilapia elegans [18].
Two types of endochondral ossification have been described in zebrafish. Type I endochondral ossification is identical to other vertebrates and is associated by formation of spongy bone. Type II lacks zone of calcification and a degradation zone and osteoblasts instead, a compact layer of chondroclasts is detected [5]. In the current study, the ribs of silver carp exhibit typical mode of endochondral ossification to type II. Vertebrae develops through endochondral bone formation of primary cartilaginous models [19]. Two forms of bone have been identified in quail embryos during growth of femur and tibia. The first form develops the physeal growth cartilage with no signs of endochondral bone formation in embryonic and juvenile stages, while, endochondral bone appears with progression of the age particularly form the tenth day of postnatal development [20]. However, it is common that physeal growth plate is associated with endochondral bone formation in mammals [21-25].
It is concluded that the ventral ribs of the silver carp grow by perichondral bone formation in addition to the linear expansion of the growth cartilage.
REFERENCES
- Boglione C, Gavaia P, Koumoundouros G and Gisbert E et al. (2013). Skeletal anomalies in reared European fish larvae and juveniles. Part 1: normal and anomalous skeletogenic processes. Reviews in Aquaculture. 5(s1): S99-S120.
- SOLIMAN SA and ABD-ELHAFEEZ HH (2014). Mesenchymal cells in cartilage growth, LAP Lambert Academic Publishing. 4: 49.
- Abd-Elhafeez HH and Soliman SA. (2016). Origin of Rodlet Cells and Mapping Their Distribution in Ruby-Red-Fin Shark (Rainbow Shark) Epalzeorhynchos frenatum (Teleostei: Cyprinidae): Light, Immunohistochemistry and Ultrastructure Study. J Cytol Histol. 7: 435.
- Abd-Elhafeez HH and Soliman SA. (2017). New Description of Telocyte Sheaths in the Bovine Uterine Tube: An Immunohistochemical and Scanning Microscopic Study. Cells Tissues Organs. 203(5): 295-315.
- Weigele J and Franz-Odendaal TA. (2016). Functional bone histology of zebrafish reveals two types of endochondral ossification, different types of osteoblast clusters and a new bone type. J Anat. 229(1): 92-103.
- Conceicao L and Tandler A. (2018). Success Factors for Fish Larval Production, John Wiley & Sons.
- Soliman AS. (2017). Potential role of telocytes in differentiation of embryonic skeletal progenitor cells. SF J Stem Cell. 1: 1.
- Soliman AS. (2018). Potential Role of Telocytes in Development of Embryonic Ganglia. SF J Stem Cell. 1: 1.
- Van Leeuwen JL. (1999). A mechanical analysis of myomere shape in fish. J Exp Biol. 202(Pt 23): 3405-3414.
- Steiner L. (2000). Pennsylvania Fish & Boat Commission.
- Huysseune A and Sire JY. (1992). Development of cartilage and bone tissues of the anterior part of the mandible in cichlid fish: a light and TEM study. Anat Rec 233(3): 357- 375.
- Dong W, Hinton DE and Kullman SW. (2012). TCDD disrupts hypural skeletogenesis during medaka embryonic development. Toxicol Sci. 125(1): 91-104.
- Benjamin M. (1990). The cranial cartilages of teleosts and their classification. J Anat 169: 153-172.
- Meunier FJ and Laurin M. (2012). A microanatomical and histological study of the fin long bones of the Devonian sarcopterygian Eusthenopteron foordi. Acta Zoologica. 93(1): 88-97.
- Hall BK and Witten PE. (2007). Plasticity of and transitions between skeletal tissues in vertebrate evolution and development. In: Anderson JS, Sues HD (eds) Major Transitions in Vertebrate Evolution, Bloomington, IN, Indiana University Press.
- Detrich III HW. The Zebrafish: Disease Models and Chemical Screens, Academic Press.
- Naruse K, Tanaka M and Takeda H. (2011). Medaka: A Model for Organogenesis, Human Disease, and Evolution, Springer Science & Business Media.
- Witten PE and Huysseune A. (2009). A comparative view on mechanisms and functions of skeletal remodelling in teleost fish, with special emphasis on osteoclasts and their function. Biol Rev Camb Philos Soc. 84(2): 315-346.
- Ytteborg E, Torgersen J, Baeverfjord G and Takle H. (2012). The Atlantic Salmon (Salmo salar) Vertebra and Cellular Pathways to Vertebral Deformities. Health and Environment in Aquaculture. E. Carvalho.
- Ahmed YA, Soliman SA and Abdel-Hafez EA. (2013). Ossification of the femur and tibia of the post-hatching Japanese quail. Pak J Biol Sci. 16(17): 859-864.
- White A and Wallis G. (2001). Endochondral ossification: a delicate balance between growth and mineralisation. Curr Biol. 11(15): R589-591.
- Yeh N, Miller JP, Tripti Gaur, Terence D, et al. (2007). Cooperation between p27 and p107 during endochondral ossification suggests a genetic pathway controlled by p27 and p130. Mol Cell Biol. 27(14): 5161-5171.
- Soliman S and Emeish W. (2017). Morphological Alternations of Intraepithelial and Stromal Telocytes in Response to Salinity Challenges. bioRxiv.
- Long F and Ornitz DM. (2013). Development of the endochondral skeleton. Cold Spring Harb Perspect Biol. 5(1): a008334.
- Soliman SA and Abd-Elhafeez HH. (2016). Are C-KIT, MMP9 and Type II Collagen Positive Undifferentiated Cells Involved in Cartilage Growth? A Description of Unusual Interstitial Type of Cartilage Growth. J Cytol Histol. 7: 440.