Information Links
Related Conferences
Previous Issues Volume 7, Issue 3 - 2022
The Genetic Evolution of “Decreased Zinc” Malignancies and Their Treatment
Leslie C Costello, PhD*
Department of Oncology and Diagnostic Sciences, University of Maryland School of Dentistry; and the University of Maryland Greenebaum Comprehensive Cancer Center, Baltimore, Maryland, USA
*Corresponding author: Dr. Leslie C Costello, PhD, Distinguished University Professor, Department of Oncology and Diagnostic Sciences, University of Maryland School of Dentistry; and the University of Maryland Greenebaum Comprehensive Cancer Center, Baltimore, Maryland 21201. Email: [email protected]
Received Date: Aug 26, 2022
Published Date: Sep 22, 2022
Citation: Costello LC. (2022). The Genetic Evolution of “Decreased Zinc” Malignancies and Their Treatment. Mathews J Cancer Sci. 7(3):34.
Copyrights: Costello LC, et al. © (2022).
ABSTRACT
The development of zinc malignancies is dependent on the relationship that “The initiation of malignancy is the evolutionary genetic transformation of normal cells to “decreased zinc” malignant cells.” The reason is the zinc concentration that exists in normal cells is cytotoxic in malignant cells. That transformation applies to the initial evolution of all malignancies. There are no exceptions! The “normal zinc” malignant cells develop in accordance with Darwin’s “Natural Selection” and the “survival of the fittest”. The human prostate gland exhibits those relationships. Testosterone-dependent acinar epithelial cells are “decreased zinc” cells; and “prolactin-dependent” acinar epithelial are “normal zinc” Natural Selection/survival of the fittest cells.
Treatment with clioquinol zinc ionophore increases the transport of zinc in the “decreased zinc” testosterone-dependent malignant cells and induces cytotoxic effects. Cabergoline prolactin agonist inhibits pituitary production and induces hypoprolactinemia, which terminates prolactin-dependent malignancy.
Keywords: decreased zinc, zinc malignancy, genetic evolution, treatment
INTRODUCTION
All cells require zinc for their reproduction, proliferation, growth, and functional activities. That includes normal cells and their corresponding malignant cells. However, the relative amount of zinc required to achieve those is different; mainly due to the process of “malignancy”. The focus of this presentation is “malignancy: its origin, progression, and treatment.”
The evolution of malignancy: “The transformation of normal cells to “decrease zinc” malignant cells.”
The contemporary views of the development and progression of malignancy have been largely mistaken and/or misrepresented by contemporary oncologists and others in the medical community. The major reason is their failure to recognize the importance of zinc in the process of the evolution of malignancy; “The initiation of malignancy is the evolutionary genetic transformation of normal cells to “decreased zinc” malignant cells.” The reason is that the zinc concentration that exists in normal cells is cytotoxic in malignant cells. That transformation applies to the initial evolution of all malignancies. There are no exceptions!
Darwin and the existence of “normal zinc” malignant cells
The next issue is the origin of “normal zinc” malignant cells. Darwin’s concept addresses that issue. He stressed the related principles of “Natural Selection” and the “survival of the fittest”. It implies that all populations of cells initially exist in conformity with their environmental conditions. Over time, the cells reproduce to sustain and repopulate their genetic phenotype for future generations. That results in gene mutations that give rise to genetically new phenotypic cells. Also, some of the existing cells are subjected to changing environmental conditions, and Darwin’s concept of the “natural selection/survival of the fittest” cells results in genetically new phenotypic populations. Therefore, at some points in time, the original population of cells and organisms and the new population of cells and organisms coexist. However, the original genotypic cells or organisms will eventually become extinct. Otherwise, the planet will be overpopulated to the point of the elimination of life on Earth.
The concept of evolution and the prostate gland
In 1999 [1], Liang et al. noted “A lower 50% inhibition of cell growth (IC50) value for zinc (about 100 ng/ml) was detected in LNCaP cells, which are androgen-responsive cells, whereas androgen-independent PC-3 cells exhibited a higher IC50 for zinc (about 700 ng/ml)”...“There now exists strong evidence that the loss of a unique capability to retain high levels of zinc is an important factor in the development and progression of malignant prostate cells.” It is the first report to identify two prostate genetic phenotypes: LNCaP cells that developed as “decreased zinc” testosterone cells; and PC-3 cells that developed as “normal zinc” prolactin cells.
Under oncogenic conditions, the normal testosterone cells are transformed to “decreased zinc” malignant cells; in conformity with the concept that “the initial development of malignancy is the evolutionary transformation of normal cells to ‘decreased zinc’ malignant cells.” That applies to all malignancies. There are no exceptions.
Then the issue is the development of “normal zinc” prolactin-dependent malignant cells. They evolved in accord with Darwin’s “natural selection/survival of the fittest” cells as described above. They are genetic mutated cells derived from testosterone-dependent “decreased zinc” malignancy in order to survive under its existing environmental conditions.
The contemporary views of the implication of zinc in malignancies
Gumulec et al. in 2002 [2] presented their meta-analysis of the of the zinc levels in human tumors based on 114 case control, cohort and cross-sectional studies. Figure 1 reveals some reports of a decrease in zinc; some reports of an increase in zinc; some reports with no change in zinc; and some reports with either an increase or decrease in zinc. Their conclusion was “The association between zinc level and stage or grade of tumor has not been revealed by meta-regression.” That is a misrepresentation of the relative concentration of zinc in malignant versus normal cells.
.jpg)
Figure 1: The relative zinc levels in human tumor tissues.
Histochemistry of zinc-stained human tissue sections establishes the transformation of normal cells to “decreased zinc” malignant cells
The histochemical analysis of zinc-stained human normal and malignant tissue sections establishes the higher zinc level in normal cells compared to the significantly decreased zinc in the malignant cells for prostate, pancreas, liver, and breast (figure) [3]. Notably, the decreased zinc in breast reveals the unreliability of determining the relative zinc levels in tissue preparations (figure 1). Also, the prostate tissue section was “decreased zinc” testosterone-dependent prostate cancer. Therefore, the histochemistry confirms the relationship that malignancy results in the transformation of normal cells to “decreased zinc” malignant cells.”
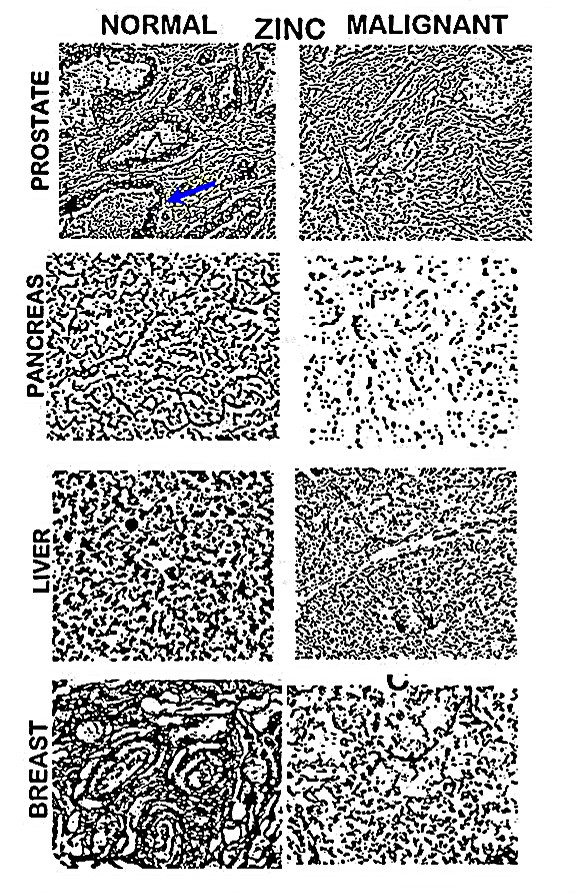
Figure 2: In situ relative zinc levels in malignant versus normal human tissue sections. Pronounced zinc exists in the normal epithelium and low zinc exists in malignancy.
ZIP-family zinc transporters: The genetic evidence for the evolution of “decreased zinc” malignancies
The issue is the genetic evidence for achieving the evolutionary transformation of normal cells to “decreased zinc” malignant cells. The zinc in blood plasma is not freely permeable across the cell membrane. Therefore, all normal mammalian cells contain a ZIP-family zinc-uptake transporter (SLC39A).
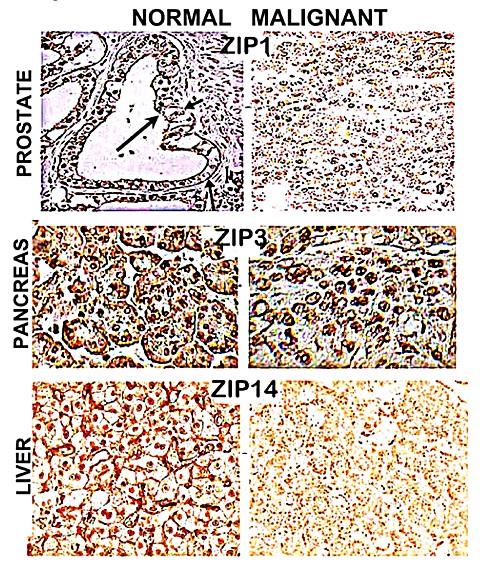
Figure 3: Relative ZIP transporter expression in malignant versus normal human prostate, liver, and pancreatic tissue sections. Arrow points to prostate acinar epithelium.
Figure 3 shows the prostate ZIP1, the pancreatic ZIP3, and the liver ZIP10 localization in the cell membrane, which transports zinc from the blood plasma into the cells [3]. It also shows the downregulation of the ZIP transporters in the malignant cells. That is the genetic evidence for the relationship: “The initial evolution is the genetic transformation of normal cells to “decreased zinc” malignant cells.” It is the dogma for all “decreased zinc” malignancies. There is no exception!
Do “increased zinc” malignancies exist?
That is a reasonable question. The answer is “No”. Despite the many reports regarding zinc and cancers, no published histochemical study exists that purportedly demonstrates an increased zinc in malignancy versus the normal cells. In the hundreds of zinc-stained prostate, pancreas, liver, and breast tissue microarrays, Costello and Franklin [3] never observed a case that exhibited an increased zinc in malignancy.
In addition, Vallee and Falchuk (pioneers and leaders regarding zinc relationships) state [4] “Clearly, a metal that is known to be essential to the inheritance of the genetic endowment and the induction of development, growth, and differentiation could not easily be intended to be deleterious to the perpetuation and evolution of the species. Instead, one would expect zinc to be regulated carefully to ensure the preservation and continuity of life. In fact, zinc is the only pre-, post-, and transitional element that has proven to be essentially nontoxic. It is neither carcinogenic, mutagenic, nor teratogenic.”
Clioquinol zinc ionophore: The treatment for patients with “decreased zinc” malignancies
Advanced testosterone-dependent prostate cancer is the major “decreased zinc” malignancy in men. Generally, its treatment has been androgen-suppression to decrease the plasma testosterone other androgens. That attenuates the progression of malignancy; but it does not terminate the malignancy.
The “decreased zinc” status provides the expectation that a treatment to increase zinc into the ZIP-deficient cells will induce cytotoxic effects that terminate the malignancy. Clioquinol zinc ionophore exhibits the zinc formation constant logKf=7-8, provides the ability to bind to 80% of the plasma ZnLigands (figure 4) [5]. The efficacy of the treatment can be established by the expression of PSA (Prostate-Specific Antigen) and/or PSMA (Prostate Specific-Membrane Antigen) prior to and 6 weeks following clioquinol treatment; i.e., the absence of PSA/PMSA-detected malignant cells .
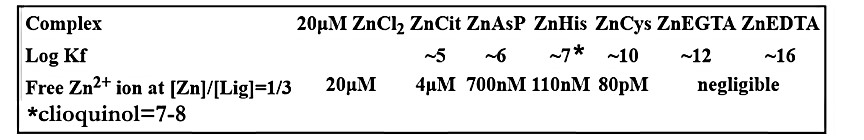
Figure 4: Clioquinol zinc-binding formation logkKf=7-8 will bind to about 80% of the zinc-exchangeable ligands in the blood plasma to produce ZnClioquinol.
The identification of prolactin-dependent prostate cancer; and its treatment
However, “androgen-independent” prostate malignancy often persists. Although differing views exist regarding the origin of those malignant cells; they have the common theme that “androgen-independent” cells represent a sub-population of the androgen-treated “advanced androgen-dependent” malignancy. A process for that transformation has not been established.
Instead, Costello and Franklin in 2002 [6] identified the existence of testosterone-dependent acinar epithelial cells and prolactin-dependent acinar epithelial cells. Their receptors and downstream signaling pathways were established. Under oncogenic conditions, testosterone-dependent prostate cancer develops, and prolactin-dependent prostate cancer develops. They are separate malignancies in which either or both can exist in patients.
Prolactin-dependent prostate cancer malignancy poses the problem that it does not express any identifying biomarkers. Its progressing malignancy is identified by the death of the patient. For that reason, I refer to advanced prolactin-dependent prostate cancer as the “silent killer” of prostate cancer patients.
That outcome must be avoided. It requires an MRI for all PSMA-negative prostate cancer patients. If the MRI detects malignancy, the patient should be treated with cabergoline. It is employed for patients with hyperprolactinemia due to a prolactin-producing pituitary adenoma. Cabergoline inhibits that production and secretion; so that the hyperprolactinemia is decreased; and the patient exhibits hypoprolactinemia. In the absence of sufficient plasma prolactin, the prolactin-dependent prostate cancer malignancy is terminated.
However, that expected outcome must be confirmed. Following the 6 weeks treatment with cabergoline, another MRI is required. If it is negative, the PSMA/MRI-negative patient is cured of testosterone-dependent prostate cancer and prolactin-dependent prostate cancer. If the MRI is positive and the plasma prolactin is decreased by greater than 80%; the patient likely has a different phenotypic cancer.
CONCLUSIONS
The relationships above are represented in the 2019 Costello et al. case report [7] and the 2022 Costello case report [8]. The patient in the 2019 report presented with advanced “decreased zinc” testosterone-dependent prostate cancer. He was first treated with clioquinol, which terminated that malignancy. Then, the PSMA patient was treated with cabergoline. An MRI was negative, and prolactin-dependent malignancy was eliminated. The 2022 report is a patient who presented with advanced “decreased zinc” testosterone-dependent prostate cancer. He was treated with clioquinol, which terminated that malignancy.
For the first time, urologists and oncologists have appropriate regimens for the diagnosis and treatment of their patients. Then, most of the estimated 280,000 men in the U.S. and the 5.3 million men worldwide, who die of cancer every year, will no longer face impending death due to cancer.
ACKNOWLEDGMENTS
L.C.C. declares that he has no conflicting interests.
The cited studies of L.C.C. were supported, in part, by NCI grant DK42839.
REFERENCES
- Liang JY, Liu YY, Zou J, Franklin RB, Costello LC, Feng P. (1999). Inhibitory effect of zinc on human prostatic carcinoma cell growth. Prostate. 40(3):200-207.
- Gumulec J, Masarik M, Adam V, Eckschlager T, Provaznik I, Kizek R. (2014). Serum and tissue zinc in epithelial malignancies: a meta-analysis. PLoS One. 9(6):e99790.
- Costello LC, Franklin RB. (2017). Decreased zinc in the development and progression of malignancy: an important common relationship and potential for prevention and treatment of carcinomas. Expert Opin Ther Targets. 21(1):51-66.
- Vallee BL, Falchuk KH. (1993). The biochemical basis of zinc physiology. Physiol Rev. 73(1):79-118.
- Costello LC, Fenselau CC, Franklin RB. (2011). Evidence for operation of the direct zinc ligand exchange mechanism for trafficking, transport, and reactivity of zinc in mammalian cells. J Inorg Biochem. 105(5):589-599.
- Costello LC, Franklin RB. (2002). Testosterone and prolactin regulation of metabolic genes and citrate metabolism of prostate epithelial cells. Horm Metab Res. 34(8):417-424.
- Costello LC, Franklin RB, Yu GW. (2019). A novel patient case report to show the successful termination of untreatable androgen-independent prostate cancer: Treatment with Cabergoline (Dopamine agonist). Mathews J Case Rep. 4(1):42.
- Costello LC. (2020). Clioquinol zinc ionophore for terminating advanced “decreased zinc” testosterone-dependent prostate cancer: A case report. Mathews J Case Rep. 7(2):73.