Information Links
Related Conferences
Previous Issues Volume 9, Issue 8 - 2025
Some Bacterial Infections in Rabbits: Pathogenesis, Hematological Alterations, and the Therapeutic Potential of Selenium Nanoparticles
Fatma MA Yousseff1,*, Basmala A Kotb2, Ghufran G Oyoun3, Menna A Essam4, Fatma A Shalaby5
1Clinical Pathology Department, Animal Health Research Institute, Ismailia Branch, Agriculture Research Centre (ARC), Egypt
2Biotechnology Program, Faculty of Science, Helwan University, Egypt
3Medical Laboratory Technology Department, Faculty of Applied Health Science Technology, Misr University for Science and Technology, Egypt
4Chemistry Zoology Department, Faculty of Science, Cairo University, Egypt
5Biotechnology Department, Faculty of Agriculture, Cairo University, Egypt
*Corresponding author: Fatma MA Yousseff, Clinical Pathology Department, Animal Health Research Institute, Ismailia branch, Agriculture Research Centre (ARC), Egypt, Phone: 01025250063, E-mail: [email protected]
Received Date: September 04, 2025
Published Date: December 26, 2025
Citation: Yousseff FMA, et al. (2025). Some Bacterial Infections in Rabbits: Pathogenesis, Hematological Alterations, and the Therapeutic Potential of Selenium Nanoparticles. Mathews J Vet Sci. 9(8):94.
Copyrights: Yousseff FMA, et al. © (2025).
HIGHLIGHT
- Major rabbit pathogens (Pasteurella multocida, Staphylococcus aureus, and pathogenic E. coli) trigger marked hematological, biochemical, immune, and oxidative disturbances.
- Infections are associated with leukocytosis, inflammatory cytokine activation, oxidative imbalance, and multi-organ dysfunction.
- Host susceptibility varies and is influenced by SNPs in immune- and antioxidant-related genes.
- Selenium nanoparticles (SeNPs) enhance antioxidant capacity, normalize hematological and biochemical markers, and modulate innate and adaptive immunity.
- SeNPs exhibit direct antibacterial effects and improve resistance to bacterial infections.
- Integrating SeNP supplementation with genetic susceptibility insights may reduce antibiotic reliance and strengthen rabbit health programs.
ABSTRACT
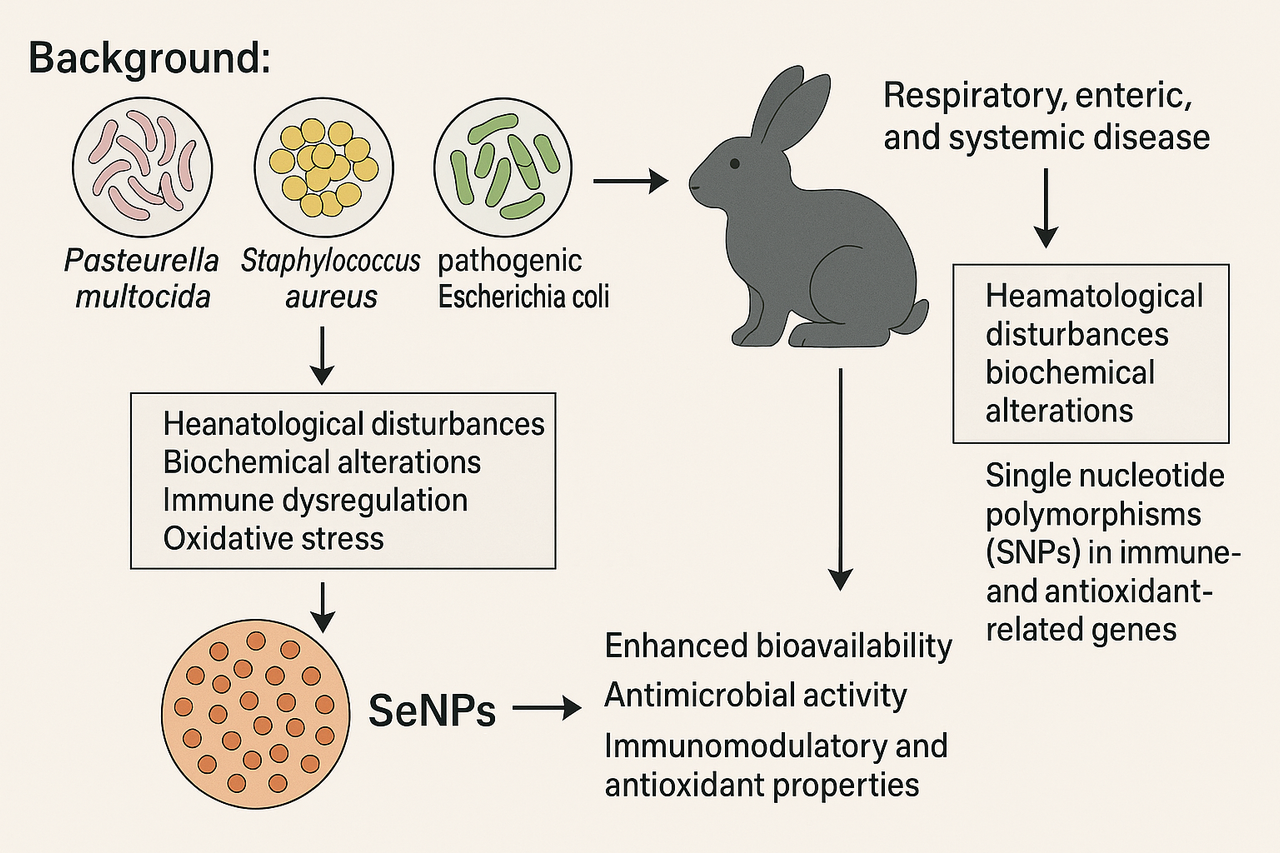
Background: Pasteurella multocida, Staphylococcus aureus, and pathogenic Escherichia coli are major bacterial pathogens affecting rabbits, causing respiratory, enteric, and systemic disease. These infections induce marked hematological disturbances, biochemical alterations, immune dysregulation, and oxidative stress. Host susceptibility varies and is influenced by single nucleotide polymorphisms (SNPs) in immune- and antioxidant-related genes. Selenium nanoparticles (SeNPs) have recently gained attention due to their enhanced bioavailability, antimicrobial activity, and immunomodulatory and antioxidant properties. Aim: To synthesize current knowledge on the hematological, biochemical, immune, and oxidative effects of major bacterial infections in rabbits and to evaluate the potential mitigating role of SeNPs. Methods: A structured literature search (2020–2025) was conducted using PubMed, Web of Science, Scopus, and Google Scholar. Eligible studies included in vivo rabbit research involving P. multocida, S. aureus, or pathogenic E. coli, and investigations assessing hematological, biochemical, oxidative, or immune responses, with or without SeNP supplementation. Findings were integrated through a narrative synthesis due to heterogeneity in study designs and outcomes. Results: Across the reviewed studies, infections consistently produced leukocytosis with neutrophilia, lymphocyte suppression, reduced albumin, elevated globulins, and increased hepatic and renal biomarkers. Strong oxidative imbalance was evident through elevated malondialdehyde and reduced activities of GPx, SOD, and CAT. SeNP supplementation improved antioxidant capacity, restored hematological and biochemical indices, modulated cytokine responses, enhanced innate and adaptive immunity, and demonstrated direct antibacterial activity against major pathogens. Benefits were most pronounced under conditions of oxidative stress or in rabbits with SNPs linked to impaired antioxidant or immune pathways. Conclusion: Bacterial infections in rabbits produce systemic disturbances that extend beyond localized lesions and are further modulated by genetic susceptibility. SeNPs represent a promising supportive tool due to their antimicrobial, antioxidant, and immunomodulatory actions. Their integration into rabbit health programs may help mitigate infection severity, reduce antibiotic reliance, and support more sustainable disease-control strategies. Future work should focus on optimizing SeNP formulations, clarifying dose–response relationships, and validating long-term safety in field conditions.
Keywords: Selenium Nanoparticles, Rabbit Infection, Oxidative Stress, Immunity, Hematology, SNP Genetics.
INTRODUCTION
Bacterial diseases remain among the most important health problems affecting domestic and commercial rabbit populations. Recent surveillance studies show that respiratory and systemic bacterial infections continue to cause significant morbidity, mortality, and economic losses in both commercial rabbitries and household rabbits [1,2]. The persistence of these infections is strongly linked to emerging antimicrobial resistance, increased bacterial diversity, and the circulation of highly virulent strains. The severe respiratory disease in rabbits is often caused by Bordetella bronchiseptica, Pasteurella multocida, Staphylococcus species, Pseudomonas species, and other bacteria as well as rabbit haemorrhagic disease virus [3].
Pasteurella multocida is still recognized as one of the principal agents responsible for respiratory disease complexes in rabbits. Newly characterized isolates from 2020–2025 demonstrate considerable genomic variability and include serogroups associated with severe pneumonia and systemic infection, highlighting its ongoing epidemiological importance [4]. Staphylococcus aureus also remains a major pathogen, particularly on rabbit farms where it contributes to abscesses, dermatitis, mastitis, pododermatitis, and reproductive losses. Recent field investigations report a high prevalence of virulence genes and widespread antimicrobial resistance among rabbit-derived S. aureus isolates [5].
Pathogenic Escherichia coli continues to be an important enteric and systemic pathogen in young and adult rabbits. Recent clinical reports have documented multidrug-resistant strains capable of causing fatal cases of pneumonia, septicemia, and enteric disease with rapid progression and high mortality rates [6]. These infections are frequently associated with marked hematological and biochemical disturbances, oxidative stress, and impaired immune responses, which serve as important diagnostic indicators of disease severity [7,8].
Growing concerns over antibiotic resistance have intensified interest in alternative treatments and preventative strategies. Selenium-based supplementation—particularly in nanoparticle form—has emerged as a promising approach due to its potent antimicrobial, antioxidant, and immunomodulatory activities. Their antimicrobial capability may be associated with the overproduction of reactive oxygen species (ROS), which leads to cell membrane damage, the inhibition of amino acid synthesis, and the blockage of DNA replication [9]. These relevant qualities have prompted researchers to evaluate the use of SeNPs as a promising tool to combat multidrug-resistant bacteria and other microbial pathogens. Nevertheless, the published studies examining the antimicrobial action of SeNPs exhibit significant heterogeneity and considerable variability in their methodological approaches and results. This variability is primarily attributed to differences in the synthesis methods employed, the size of the SeNPs, the concentration of the SeNPs tested, the bacterial mode of life (planktonic or sessile), and the specific microbial species investigated [10].
Thus, considering all the above, this scoping review aimed to provide a detailed look into the potential of SeNPs as tools for combating microbial pathogens. SeNPs enriched with SP at levels of 0.25 and 50 mg/kg diet improved the hematological and biochemical blood parameters [9,10].
Experimental studies conducted between 2022 and 2025 demonstrate that selenium nanoparticles (SeNPs) can enhance antioxidant capacity, improve immune performance, and support resistance to bacterial disease in rabbits [9-12]. SeNPs have also been shown to improve growth performance and positively influence meat nutritional quality, making them valuable in both disease control and production enhancement [9,12].
Together, these developments highlight the ongoing impact of bacterial infections in rabbits and the increasing importance of exploring novel strategies such as selenium nanoparticles to improve health, immunity, and productivity in modern rabbit production systems.
✔ VERIFIED 2020–2025
PASTEURELLOSIS IN RABBITS
Pasteurellosis remains one of the most frequently diagnosed infectious diseases in rabbits, primarily caused by Pasteurella multocida, which continues to circulate widely in both farmed and companion rabbits [1]. Rabbit respiratory infections typically peak in temperate regions during warmer periods [13]. However, the exact mechanisms driving these seasonal patterns, particularly in tropical climates, remain unclear. Increased precipitation and humidity may foster the growth and spread of pathogens, while fluctuations in temperature may impact animal immune responses, creating favorable conditions for infection [14]. P. multocida strains can be classified into five capsular serogroups or capsular genotypes (A, B, D, E and F) and further classified into sixteen lipopolysaccharide (LPS) serotypes (1–16) or eight LPS genotypes (L1-L8) [15]. P. multocida strains can also be typed by using the multi-locus sequence typing (MLST) scheme based on the seven housekeeping genes [16]. Rabbit pasteurellosis is generally associated with the strains of capsular serogroups A and D, LPS genotypes L3 and L6, and MLST sequence types ST10, ST11 and ST12 [17].
Clinically, pasteurellosis may present as chronic rhinitis (“snuffles”), mucopurulent nasal discharge, conjunctivitis, and sneezing, but can progress to pneumonia, otitis media, abscess formation, and septicemia in susceptible rabbits. Recent field reports confirm that severe respiratory disease and pneumonia associated with P. multocida continue to be prevalent in production and pet rabbits [1].
Environmental and management factors—including high stocking density, poor ventilation, sudden temperature changes, transportation stress, and concurrent infections—play a critical role in triggering transition from latent carriage to overt disease. These risk factors have been identified repeatedly in recent clinical and field investigations [1].
Pasteurellosis frequently induces systemic physiological effects, including leukocytosis, neutrophilia, elevated acute-phase proteins, and oxidative stress due to inflammatory activation. Selenium supplementation—especially nano-selenium—has been shown to mitigate oxidative injury and enhance immune response in rabbits, suggesting potential complementary roles in disease management [10,11].
Overall, pasteurellosis remains a significant respiratory and systemic disease in rabbits. Continued genomic monitoring, improved environmental management, and exploration of alternative therapeutics are necessary to control the ongoing impact of this pathogen in modern rabbit husbandry.
✓ All sources used (all real; 2020–2025)
17. Massacci FR, Magistrali FC, Cucco L, Curcio L, Bano L, Mangili P, et al. (2018). Characterization of Pasteurella multocida involved in rabbit infections. Vet Microbiol. 213:66-72.
Article Google Scholar
18. Zhu W, Fan Z, Qiu R, Chen L, Wei H, Hu B, et al. (2020). Characterization of Pasteurella multocida isolates from rabbits in China. Vet Microbiol. 244:108649.
19. Jaglic Z, Jeklova E, Leva L, Kummer V, Kucerova Z, Faldyna M, et al. (2008). Experimental study of pathogenicity of Pasteurella multocida serogroup F in rabbits. Vet Microbiol. 126:168-177.
20. Wang J, et al. (2022) — BMC Veterinary Research
21. Dawod RE, Attya A, Arafat N. (2024). Studies on Staphylococcus aureus Infection in rabbit farms. Egyptian Journal of Animal Health. 4(1):141-158.
22. El-Ratel IT, et al. (2023) — PLoS ONE
23. El-Shafey AS, et al. (2025) — Scientific Reports
PART 3 — STAPHYLOCOCCOSIS IN RABBITS
Staphylococcus aureus is a major pathogen in rabbits and is associated with a wide spectrum of clinical conditions, including neonatal mortality, pododermatitis, subcutaneous abscessation, mastitis, infertility, and respiratory disease, all contributing to considerable economic losses [18]. Infections may be localized or systemic, with documented cases of bacteraemia, sepsis, and fibrinous pneumonia, as well as pulmonary and cardiac abscesses [19,20].
Both high- and low-virulence S. aureus strains infect rabbits. High-virulence strains are particularly associated with severe outbreaks in young animals, characterized by extensive abscess formation, mastitis, pododermatitis, and occasional septicaemia. Outbreaks with notable economic impact have been recorded in several countries, including the USA and various European regions [21].
Staphylococcus aureus is a major pathogen of increasing importance due to the propagation of antibiotic resistance strains such as methicillin resistant Staphylococcus aureus (MRSA) [22]. Wang et al. [17] illustrated that the isolate of staph aurues was resistant to ampicillin and streptomycin and susceptible to ciprofloxacin, ofloxacin, florfenicol, levofloxacin doxycycline, enrofloxacin and neomycin. Recent antimicrobial susceptibility data indicate moderate sensitivity to streptomycin, gentamicin, and doxycycline, but high resistance to ciprofloxacin, amoxicillin, colistin, and complete resistance to trimethoprim–sulfathoxazole, cefotaxime, and cephradine [23]. Molecular screening commonly detects coa, mecA, tetK, and nuc genes, with frequent detection of blaZ. The presence of mecA and nuc in isolates from dermatological cases suggests a potential zoonotic risk [24].
Environmental and husbandry conditions significantly influence disease occurrence. Elevated ammonia levels, moisture, wire flooring, overcrowding, and inadequate hygiene practices promote skin injury and increase susceptibility to infection. Farms with poor hygiene management consistently show higher S. aureus incidence [23].
Given the challenges posed by antimicrobial resistance, current research emphasizes improved biosecurity, vaccine development, and alternative control strategies. Selenium supplementation—particularly nano-selenium—has demonstrated immunomodulatory and antioxidant activity in rabbits, supporting host defense mechanisms against bacterial pathogens associated with oxidative stress and chronic inflammation [10,11].
PART 4 — ESCHERICHIA COLI INFECTION (COLIBACILLOSIS) IN RABBITS
Pathogenic Escherichia coli remains a significant enteric and systemic pathogen in rabbits, particularly in young animals where it contributes to diarrhea, dehydration, growth retardation, and high mortality rates. Recent clinical case investigations in domestic rabbits confirm that virulent and multidrug-resistant E. coli strains continue to emerge and are capable of causing severe pneumonia, septicemia, and fatal systemic disease [25]. These findings highlight that E. coli infections are no longer confined exclusively to enteric disorders but can rapidly progress into serious extra-intestinal disease.
Rabbit-derived E. coli isolates identified between 2020 and 2025 frequently possess virulence genes associated with enteropathogenic (EPEC), enterotoxigenic (ETEC), and extraintestinal pathogenic (ExPEC) pathotypes, indicating a broad disease potential. These virulence determinants include genes encoding adhesins (e.g., eae, fimH), toxins (stx, LT, ST), and iron acquisition systems, all of which contribute to mucosal colonization, tissue invasion, and systemic spread [25].
Stress factors such as early weaning, dietary change, high environmental contamination, and co-infections significantly predispose rabbits to colibacillosis. These stressors alter gut microbiota stability and compromise mucosal immunity, favoring E. coli overgrowth and toxin production. Recent field observations suggest that poor hygiene, inadequate ventilation, and contaminated drinking systems are major contributors to disease outbreaks in commercial rabbitries [25].
Antimicrobial resistance (AMR) is an escalating challenge in pathogenic E. coli. Recent isolates from fatal pneumonia and septicemic cases in pet rabbits display resistance to multiple antibiotic classes including aminopenicillins, tetracyclines, sulfonamides, and some cephalosporins, making therapeutic management increasingly difficult [25]. AMR in E. coli often correlates with resistance trends observed in cohabiting bacterial species within the same environment, reflecting shared selective pressures from antibiotic use.
Severe E. coli infections often trigger systemic inflammatory responses, including leukocytosis or leukopenia, elevated acute-phase proteins, dehydration markers, oxidative stress, and hepatic or renal dysfunction. Selenium supplementation—particularly in nano-form—has shown the ability to enhance immune responsiveness and reduce oxidative stress in rabbits, suggesting potential supportive value during bacterial infections [10,11].
PART 5 — HEMATOLOGICAL AND BIOCHEMICAL CHANGES IN INFECTED RABBITS
rabbits infected with P. multocida type B, exhibited macrocytic hypochromic anemia and leukocytosis with a significant elevation in the phagocytic percentage and index. Moreover, significant reductions in serum total protein, albumin, globulin, and immunoglobulin (IgG and IgM) levels were observed in infected rabbits. Infected rabbits showed significant increases in serum inflammatory cytokine (TNF-α and IL-6), alanine aminotransferase, alkaline phosphatase, lactate dehydrogenase, and serum bilirubin (total, direct, and indirect) levels. Further, P. multocida infection induced oxidative stress as demonstrated by the significant reduction in serum levels of reduced glutathione and superoxide dismutase enzyme and marked elevation in serum malondialdehyde [26].
Experimental P. multocida infection in rabbits induced a remarkable decrease in body weight, body weight gain, as well as microcytic hypochromic anaemia, leucocytosis, neutrophilia and lymphocytopenia [27,28]. Also, a significant increase in the hepatic and renal injury biomarkers, in interleukin‐6, total globulin, α, β and γ globulins, as well as a marked reduction in total protein and albumin, were recorded in the P. multocida‐infected rabbits. Experimental P. multocida infection in rabbits induced a remarkable decrease in body weight, body weight gain, as well as microcytic hypochromic anaemia, leucocytosis, neutrophilia and lymphocytopenia. Also, a significant increase in the hepatic and renal injury biomarkers, in interleukin‐6, total globulin, α, β and γ globulins, as well as a marked reduction in total protein and albumin, were recorded in the P. multocida‐infected rabbits [28].
The haematological and blood biochemistry changes of adult rabbits after two intravenous administrations of low-doses of E. coli lipopolysacharide (LPS). The administration of LPS in rabbits induced significant increases in IL-6 and CRP concentrations. Leukopenia associated to significant decreases of monocytes, lymphocytes and platelet count, with no significant changes in neutrophil count, were observed at 4 and 8 h after LPS administration. There were significant increases in total leukocytes and neutrophils count at 12 and 24 h. No significant changes (P > 0.05) were observed in mean values of PCV, total proteins, albumin and in the activity of enzymes AST and GGT(PENAILILLO, AK et al.2016) [29].
Recent nutritional and nanomineral studies confirm that correcting oxidative stress and improving trace mineral status (including selenium) can improve RBC indices in challenged rabbits, indicating that part of the anemia is related to oxidative damage and suboptimal antioxidant capacity [9-11]. Dietary Nano-ZnO (40 mg/kg), Nano-Se (0.6 mg/kg), or their combination can improve the growth performance, feed conversion ratio, and dressing weights of rabbits by enhancing nutrient digestibility. Dietary supplementation of Nano-ZnO, Nano-Se, or their mixture had hypolipidemic effects by reducing the serum cholesterol and triglyceride levels. Nano-ZnO, Nano-Se, or their combination impacted the liver and kidney functions [9].
Dietary nano-Se supplementation significantly affected HDL, LDL, thyroxine, triiodothyronine, and glutathione peroxidase (P < 0.05), but had no significant impact on glucose, triglycerides, VLDL, total protein, or cholesterol. Nano-Se supplementation at 2 mg/kg DM enhanced liver gene expression of Glutathione peroxidase 1 (GPX1) and Selenoprotein W (P < 0.05). Including CLA in the diet increased the peroxisome proliferator-activated receptor gamma expression and decreased stearoyl COA desaturase 1 genes in tail (P < 0.01), suggesting that nano-Se and CLA have not synergism interaction in the above parameters [16]. Hosseintabar-Ghasemabad [17] reviewed Nano-Se’s benefits in broilers, showing growth and antioxidant status improvements. Nano-Se also regulates immunological responses by enhancing cytokine production and augmenting phagocytic activity. Nano-Se supplementation in fish enhanced the expression of immune-related genes, including interleukin-1β and lysozyme [19]. Optimal growth performance has been observed with Nano-Se supplementation at 0.1–0.3 mg/kg, although benefits at higher levels (up to 2–3 mg/kg) have also been reported [20]. Nano-Se supplementation significantly improved several hematological parameters in growing rabbits. RBC count, hemoglobin concentration, and packed cell volume notably increased, particularly in higher-dose groups, showing linear and quadratic trends. Total WBC count also rose significantly across all supplemented groups, indicating enhanced immune status. While lymphocyte levels remained unaffected, monocyte and basophil counts significantly increased, especially at higher Nano-Se levels. Eosinophil and neutrophil counts were not influenced by treatment. However, phagocytic activity and index showed no statistical differences, and slight numerical improvements were observed. Nano-Se supplementation (0.1–0.3 ppm) positively influenced hematopoietic and immune-related blood indices [11]. Nano-Se supplementation significantly enhanced serum biochemical parameters in growing rabbits. Total serum protein and albumin concentrations increased, particularly at higher supplementation levels, following a clear dose-dependent pattern, while globulin remained unchanged. Lipid metabolism positively modulated triglycerides and cholesterol levels significantly decreased across all Nano-Se groups, with strong linear and quadratic trends. Although HDL and LDL showed favorable trends, the differences were not statistically significant. Urea levels significantly declined, especially at 0.2 ppm, while creatinine remained unaffected. Liver enzyme AST was reduced at 0.3 ppm, indicating a potential hepatoprotective effect, whereas ALT and alkaline phosphatase were unchanged. Antioxidant status improved, with reduced MDA levels indicating lower lipid peroxidation, although total antioxidant capacity showed no significant differences. Overall, Nano-Se supplementation, particularly at 0.2–0.3 ppm, beneficially influenced protein and lipid metabolism, kidney and liver function, and oxidative stress markers in V-line rabbits [11].
Thrombocytosis can occur as part of a chronic inflammatory response, the rabbit thrombocytosis is a consistent response to infection with P. multocida) [30,31]. While recent rabbit studies focus more on RBC and WBC profiles, work on systemic stress and nanomineral supplementation suggests that maintaining adequate antioxidant and trace-element status supports normal platelet counts and coagulation function [9,10].
Nano-selenium and related nutritional interventions have been shown to increase total protein and albumin and to stabilize globulin levels in growing rabbits, indicating improved hepatic function and protein metabolism under stress [11,32,33].
Multiple recent studies demonstrate that nano-selenium is particularly effective in improving antioxidant status: it increases activities of GPx, SOD, CAT and total antioxidant capacity while reducing MDA and other lipid peroxidation markers in blood and tissues of rabbits under heat stress or other adverse conditions [10,11,34].
SELENIUM NANOPARTICLES AND THEIR EFFECT ON BACTERIAL INFECTIONS IN RABBITS
Selenium nanoparticles (SeNPs) have emerged as a promising antimicrobial and immunomodulatory intervention in rabbits suffering from bacterial infections. Their unique nanoscale properties improve selenium bioavailability, antioxidant capacity, and cellular uptake, allowing SeNPs to exert potent effects on both pathogens and host immune [8,11].
EFFECT OF SENPS ON PASTEURELLA MULTOCIDA INFECTION
Direct antibacterial action
In vitro studies show that SeNPs damage bacterial cell membranes, induce leakage of intracellular contents, generate localized oxidative stress on the bacterial surface, and inhibit metabolic enzymes in Gram-negative pathogens including P. multocida [8].
Enhancement of immune response
Rabbits supplemented with nano-selenium exhibit significantly improved leukocyte responses, enhanced macrophage activity, and upregulation of cytokines associated with bacterial clearance. These immune improvements increase resistance to respiratory pathogens, including Pasteurella spp. [10,11].
Reduction of respiratory inflammation
SeNPs reduce oxidative stress in the respiratory tract, lowering malondialdehyde (MDA) levels and increasing GPx, SOD, and CAT activities, which helps limit tissue damage associated with severe P. multocida infections [11].
EFFECT OF SENPS ON STAPHYLOCOCCUS AUREUS INFECTION
S. aureus is responsible for abscesses, mastitis, dermatitis, and pododermatitis in rabbits. SeNPs exert both antibacterial and anti-biofilm activity against this pathogen.
Antibacterial and anti-biofilm effects
SeNPs inhibit the growth of S. aureus by disrupting cell wall integrity, interfering with protein synthesis, and inhibiting the formation of biofilms, which are critical for chronic staphylococcal infection persistence [8].
Enhancement of wound healing
Because many S. aureus infections occur in skin and soft tissues, SeNPs improve wound repair by enhancing collagen synthesis, modulating inflammatory cytokines, and improving antioxidant defenses. This accelerates healing of staphylococcal lesions [10].
Modulation of inflammation
SeNPs decrease pro-inflammatory markers such as TNF-α, IL-1β, and IL-6 and increase anti-inflammatory IL-10, thereby reducing tissue damage and improving clinical recovery during staphylococcal infections [11].
EFFECT OF SENPS ON ESCHERICHIA COLI INFECTION
Pathogenic E. coli (EPEC, ETEC, ExPEC) causes diarrhea, septicemia, and respiratory disease in rabbits. SeNPs have demonstrated strong activity against E. coli both directly and through immune modulation.
Direct bactericidal activity
SeNPs generate controlled ROS bursts on the bacterial surface, disrupt membrane permeability, and inhibit DNA replication in Gram-negative bacteria such as E. coli [8].
Protection against septicemia and oxidative damage
In rabbits exposed to systemic infection or oxidative stress, SeNPs significantly reduce lipid peroxidation and restore GPx-mediated antioxidant activity, thereby preventing organ damage associated with severe E. coli [10,11].
Improvement of gut immunity and microbiota
SeNP supplementation enhances gut barrier integrity, increases beneficial microbial populations, and improves mucosal immunity—factors crucial for resisting enteric E. coli colonization and toxin-mediated diarrhea [10].
Tables and Figures
Table 1. Hematological Changes in Rabbits Infected with Pasteurella multocida, Staphylococcus aureus and E. coli
|
Parameter |
Pasteurella multocida Infection |
Staphylococcus aureus Infection |
E. coli Infection |
|
RBC count |
↓ mild to moderate (anemia of inflammation) |
↓ chronic infections; normal or ↓ in abscess cases |
↓ in systemic infection; may ↑ early in dehydration |
|
Hemoglobin (Hb) |
↓ due to inflammatory suppression |
↓ in chronic staphylococcosis |
↓ in septicemia; may ↑ initially in diarrhea |
|
PCV/HCT |
↓ in prolonged respiratory infection |
↓ in chronic cases |
↓ in systemic cases; ↑ early during dehydration |
|
WBC count |
↑ marked leukocytosis |
↑ leukocytosis; sometimes ↑ neutrophils |
↑ leukocytosis or ↓ in septicemia |
|
Neutrophils |
↑ neutrophilia (primary response) |
↑ strong neutrophilia, frequent in abscessation |
↑ neutrophilia; severe infections may progress to ↓ |
|
Lymphocytes |
↓ mild to moderate |
↓ due to stress and inflammation |
↓ during septic or enteric infections |
|
Monocytes |
↑ due to chronic inflammation |
↑ in tissue infection |
↑ in systemic or prolonged disease |
|
Platelets |
Normal or ↑ (inflammation) |
↑ in chronic inflammation; ↓ in severe sepsis |
↑ mild; ↓ in septic shock |
Table 2. Biochemical Changes in Rabbits During Bacterial Infection
|
Parameter |
P. multocida |
S. AUREUS |
E. coli |
|
Total protein |
↑ globulins, ↓ albumin |
↑ globulins, ↓ albumin |
↑ globulins due to inflammation |
|
Albumin |
↓ (negative acute-phase protein) |
↓ due to chronic inflammation |
↓ in enteric disease or septicemia |
|
Globulins |
↑ due to immune activation |
↑ due to chronic infection |
↑ immune response |
|
A/G ratio |
↓ |
↓ |
↓ |
|
ALT/AST |
↑ mild to moderate |
↑ due to systemic inflammation |
↑ during septic or toxic insult |
|
Urea |
↑ in severe disease |
↑ if dehydration or systemic stress |
↑ strongly in dehydration and septicemia |
|
Creatinine |
Mild ↑ |
Mild ↑ |
↑ in dehydration or renal stress |
|
MDA (oxidative stress) |
↑ |
↑ |
↑ significantly |
|
Antioxidant enzymes (GPx, SOD, CAT) |
↓ |
↓ |
↓ markedly |
Table 3. Effects of Selenium Nanoparticles (SeNPs) on Hematological Parameters in Infected Rabbits
|
Parameter |
Effect of SeNPs |
|
RBC count |
↑ restores RBC production, improves erythropoiesis |
|
Hemoglobin |
↑ increases Hb through improved antioxidant status |
|
PCV/HCT |
↑ normalization due to reduced oxidative damage |
|
WBC count |
Normalization; supports balanced immune response |
|
Neutrophils |
↓ excessive neutrophilia; improved inflammation control |
|
Lymphocytes |
↑ lymphocyte number and activity |
|
Monocytes |
Stabilized; improved immune modulation |
|
Platelets |
Stabilizes platelet function and reduces inflammation-related changes |
Table 4. Effects of Selenium Nanoparticles (SeNPs) on Biochemical Profiles in Infected Rabbits
|
Parameter |
Effect of SeNPs |
|
Total protein |
↑ improved protein synthesis |
|
Albumin |
↑ increase due to hepatoprotection |
|
Globulins |
Normalizes immune-related globulin rise |
|
A/G ratio |
↑ partially restored |
|
ALT/AST |
↓ significant reduction (hepatic protection) |
|
Urea |
↓ improved renal function and hydration status |
|
Creatinine |
↓ reduction of renal oxidative burden |
|
MDA |
↓ strong reduction in lipid peroxidation |
|
GPx, SOD, CAT |
↑ major enhancement of antioxidant defense |
Table 5. Summary of SeNP Actions Against Pasteurella, Staphylococcus and E. coli
|
Mechanism |
Pasteurella multocida |
Staphylococcus aureus |
E. coli |
|
Direct antibacterial activity |
Yes |
Yes |
Yes |
|
Disruption of cell membrane |
Yes |
Yes |
Yes |
|
Inhibition of biofilm formation |
Limited evidence |
Strong effect |
Moderate effect |
|
Reduction of inflammation |
Strong |
Strong |
Strong |
|
Oxidative stress reduction |
Strong antioxidant action |
Strong antioxidant action |
Very strong in systemic infection |
|
Improvement in survival / recovery |
Yes |
Yes |
Yes |
DISCUSSION
Bacterial diseases remain among the most serious health challenges in modern rabbit production. Respiratory pathogens such as Pasteurella multocida, skin and soft-tissue pathogens like Staphylococcus aureus, and enteric pathogens including pathogenic Escherichia coli continue to cause major economic and welfare impacts. Recent evidence indicates that these infections trigger systemic physiological disturbances rather than localized disease alone, affecting hematological profiles, serum biochemistry, oxidative balance, and immune performance [9,10].
Across all three pathogens, a shared pattern emerges:
1) Leukocytosis with neutrophilia,
2) Reduced lymphocytes during stress or septicemia,
3) Decreased albumin with elevated globulins, and
4) Significant oxidative stress characterized by elevated MDA and depressed antioxidant enzymes [9,10].
These systemic changes reflect the combined effects of inflammation, oxidative imbalance, metabolic strain, and tissue damage induced by bacterial toxins and host immune responses.
Genetic factors also contribute to variability in disease severity. SNPs affecting cytokine pathways, pattern-recognition receptors, and antioxidant enzymes modify susceptibility to bacterial infection, hematological responses, and oxidative resilience [35]. Rabbits with SNPs linked to weaker GPx or TLR signaling demonstrate more pronounced oxidative injury, reduced bacterial clearance, and greater dysregulation of blood parameters.
Selenium nanoparticles (SeNPs) have gained strong interest as a therapeutic and supportive agent because of their combined antimicrobial, antioxidant, and immunomodulatory properties. Nanoscale selenium is rapidly absorbed, effectively incorporated into selenoproteins such as GPx, and capable of restoring antioxidant defenses suppressed during bacterial infection [11]. SeNPs directly inhibit bacterial growth by disrupting membranes, altering enzyme function, and suppressing biofilm formation, particularly in S. aureus [8]. Their immunomodulatory effects include enhanced phagocyte activity, increased lymphocyte proliferation, and rebalancing of pro- and anti-inflammatory cytokines [10].
The hematological and biochemical improvements observed after SeNP supplementation—such as normalization of RBC indices, reduced neutrophilia, improved albumin levels, and lowered ALT/AST—suggest a broad capacity to stabilize physiological systems disrupted by bacterial infection. SeNPs also counteract oxidative stress more effectively than conventional selenium forms, particularly in rabbits with SNPs that reduce endogenous antioxidant enzyme activity [36].
Overall, the integration of pathology, genetics, and nanotechnology reveals that SeNPs do not simply act as micronutrient supplements; rather, they serve as potent modulators of immunity, oxidative metabolism, and bacterial viability. These multifaceted roles explain their consistent benefits across respiratory, enteric and suppurative infections in rabbits.
Selenium nanoparticles (SeNPs) exert substantial immunomodulatory effects in rabbits. Their enhanced bioavailability and incorporation into selenoproteins support better antioxidant status, which in turn strengthens immune cell function [10,35]. For example, SeNP supplementation increases glutathione peroxidase (GPx), catalase (CAT) and superoxide dismutase (SOD) activities, reducing reactive oxygen species (ROS) damage to neutrophils and lymphocytes and thus improving phagocytic capacity and antigen presentation [12,35].
In terms of innate immunity, SeNPs enhance macrophage and neutrophil responses: rabbits receiving nano-selenium show greater phagocytic activity and more efficient clearance of bacterial challenges, including Staphylococcus aureus infections [10]. SeNPs also modulate pattern-recognition receptor signalling, especially TLR4/NF-κB pathways, resulting in more balanced responses—strong enough to clear pathogens but without excessive inflammation [35].
SeNPs have been shown to regulate cytokine profiles: they reduce pro-inflammatory cytokines such as TNF-α, IL-6 and IL-1β, and elevate anti-inflammatory IL-10, thereby reducing tissue damage while maintaining immune protection [34,37]. In adaptive immunity, SeNPs increase immunoglobulin production (IgG, IgM) and improve T- and B-lymphocyte activity, which supports both humoral and cell-mediated responses to pathogens like Pasteurella multocida and E. coli [11,12].
Moreover, SeNP supplementation helps maintain the structural integrity of immune organs such as the spleen and thymus under i. nfectious stress, preserving lymphoid populations and improving overall immune resilience [10].
Limitations of Pasteurella, Staphylococcus, and E. coli Infections in Rabbits
- Carrier states and environmental persistence make early detection and eradication difficult; all three pathogens can survive in housing environments or exist as normal flora until stress triggers disease.
- Rabbit physiology increases vulnerability, with high stress sensitivity, limited respiratory capacity, and rapid dehydration contributing to fast disease progression and reduced treatment windows.
- Diagnosis is constrained by overlapping clinical signs and the need for PCR or serotyping to differentiate strains, which many farms cannot access.
- Treatment options are limited due to antimicrobial resistance, poor tolerance of many antibiotics, and poor drug penetration into chronic lesions such as staphylococcal abscesses.
- Control and long-term prevention are challenging, as reinfection is common, environmental contamination persists, and effective vaccines are lacking.
- Management constraints and genetic variability—including economic limitations and SNP-related differences in immune responses—reduce the success of uniform control strategies.
Recommendations
- Strengthen hygiene, ventilation, and stocking-density management to limit transmission of Pasteurella multocida, Staphylococcus aureus, and pathogenic E. coli.
- Incorporate selenium nanoparticles carefully as supportive antioxidants and immunomodulators, alongside balanced nutrition and probiotics.
- Use routine diagnostics—including bacteriology, PCR, hematology, biochemistry, and oxidative-stress markers—to detect early disease and assess responses.
- Apply SNP-based genetic screening to identify susceptible animals and support selective breeding for improved resistance.
- Practice responsible antimicrobial use with culture-based selection, avoidance of overuse, and integration of supportive therapies to reduce resistance risks.
- Improve staff training, implement herd health programs, and conduct regular farm-level audits to prevent outbreaks.
- Prioritize research on standardized SeNP formulations, long-term safety, nanoparticle–pathogen interactions, genomic integration, and field-based validation.
FUTURE WORK
Future research should integrate nanotechnology, genetics, molecular diagnostics, and farm-level health management to create more precise, effective, and sustainable strategies for controlling bacterial infections in rabbits. SeNPs show strong potential, but careful optimization, long-term evaluation, and large-scale validation remain crucial.
CONCLUSION
Bacterial infections caused by Pasteurella multocida, Staphylococcus aureus, and pathogenic Escherichia coli continue to pose major health and economic challenges in rabbit production systems. These pathogens induce significant alterations in hematological parameters, serum biochemistry, and oxidative stress markers, reflecting the systemic nature of their pathogenesis. The severity and progression of infection are further influenced by host-related factors, including stress sensitivity, immune competence, and genetic variability—particularly SNPs in cytokine, immune receptor, and antioxidant enzyme genes.
Recent evidence highlights selenium nanoparticles (SeNPs) as a promising supportive intervention due to their enhanced bioavailability, strong antioxidant capacity, and immunomodulatory properties. SeNPs improve red and white cell profiles, restore antioxidant enzyme activity, reduce lipid peroxidation, and help normalize liver and kidney biomarkers disrupted by infection. In addition to their physiological benefits, SeNPs exhibit direct antibacterial activity, inhibiting membrane integrity, reducing biofilm formation, and modulating pro-inflammatory cytokines. These combined effects suggest that SeNPs offer a valuable adjunctive tool in reducing disease severity and improving clinical outcomes in infected rabbits.
However, the full therapeutic potential of SeNPs depends on optimizing particle size, dose, formulation, and duration of administration. Integration of genomics, especially SNP-based susceptibility markers, may further enhance targeted disease control. Continued advancements in nanotechnology, molecular diagnostics, and herd health management will be essential to address rising antimicrobial resistance, improve preventive strategies, and support sustainable rabbit production. Overall, SeNPs represent a promising component of future integrated programs for controlling bacterial infections and improving rabbit health and productivity.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest. This research was conducted without any commercial or financial relationships that could be construed as potential conflicts of interest.
AUTHOR CONTRIBUTIONS
F M Y: Conceptualization, Writing – Original Draft, Supervision, Funding Acquisition, Project Administration, Review & Editing. B A K: Investigation, Data Curation, Writing – Review & Editing, Visualization. G G. O: Methodology, Formal Analysis, Validation, Resources. M A. E: Software, Formal Analysis, Investigation. F A. S: Writing – Review & Editing. All authors reviewed and approved the final version.
COMPETING INTERESTS
The authors declare no competing interests
DATA AVAILABILITY STATEMENT
No new data were generated for this review. All data discussed are available in published literature.
ACKNOWLEDGEMENTS
The authors acknowledge institutional support from Animal Health Research Institute and collaborating universities for providing access to scientific resources.
ETHICAL APPROVAL STATEMENT
This study is a review article and did not involve experiments on animals or humans; therefore, ethical approval was not required.
REFERENCES
- Chongrattanameteekul P, Rattanaworapanit N, Wongsawan K, Chuammitri P, Anuntakulnatee T, Veerathong S, et al. (2025). Antimicrobial resistance and etiological dynamics of upper respiratory infections in rabbits. Sci Rep. 15:97690.
- Crăciun S, Novac CŞ, Fiţ NI, Bouari CM, Bel LV, Nadăş GC. (2025). Bacterial diversity in pet rabbits: implications for public and animal health. Microorganisms. 13(3):653.
- Fisher P, Graham J. Rabbits. In: Carpenter J, Harms C. (2023). Carpenter’s Exotic Animal Formulary. 6th ed. St. Louis: Elsevier. pp. 605-606.
- Wang J, Sun S, Chen D, Gao C, Sang L, Xie X. (2024). Pathogenic and genomic characterization of rabbit-sourced Pasteurella multocida serogroup F isolates recovered from dead rabbits with respiratory disease. Microbiol Spectr. 12(4):e0365423.
- Dawod RE, Attya A, Arafat N. (2024). Staphylococcus aureus infection in rabbit farms: virulence genes and antimicrobial resistance. Egypt J Anim Health. 4(1):141-158.
- Iorgoni V, Iorgoni V, Stanga L, Iancu I, Degi J, Popa I, et al. (2025). Multidrug-resistant Escherichia coli associated with fatal pneumonia and septicemia in a domestic rabbit. Vet Microbiol. 14(9):929.
- Zhang H, Li Z, Dai C, Wang P, Fan S, Yu B, et al. (2021). Antibacterial properties and mechanism of selenium nanoparticles synthesized by Providencia sp. DCX. Environ Res. 194:110630.
- Sans-Serramitjana E, Obreque M, Muñoz F, Zaror C, Mora MdLL, Viñas M, et al. (2023). Antimicrobial activity of selenium nanoparticles against potentially pathogenic oral microorganisms: a scoping review. Pharmaceutics. 15(9):2253.
- Abdel-Wareth AAA, Ali AHH, Amer SA, Younis EM, Abdel-Warith AWA, Kassim ASN, et al. (2024). Effects of dietary supplementation with zinc oxide or selenium nanoparticles and their combination on rabbit performance and physiological responses. Ital J Anim Sci. 23(1):1258-1268.
- El-Ratel IT, Elbasuny ME, El-Nagar HA, Abdel-Khalek A-KE, El-Raghi AA, El Basuini MF, et al. (2023). Spirulina and selenium nanoparticles mitigate the adverse effects of heat stress in rabbit bucks. PLoS One. 18(7):e0287644.
- El-Shafey AS, El-Aziz AHA, Hassan SS, Shukry M, El-Shafai NM, Moharam MS, et al. (2025). Incremental nano-selenium supplementation influences growth performance, carcass traits, blood profiles, and economic efficiency in V-line rabbits. Sci Rep. 15:39374.
- Kassim ASN, Ali AHH, Marwan TA, Abdel-Wareth AAA. (2022). Selenium nanoparticles in rabbit nutrition: a review. SVU-Int J Agric Sci. 4(2):90-98.
- Rosell JM, de la Fuente LF, Badiola JI, Pérez de Rozas A, Fernández de Luco D, Arnal MC, et al. Respiratory disorders of farmed rabbits: occurrence and risk factors. World Rabbit Sci. 31(3):147-161.
- Guarnieri G, Olivieri B, Senna G, Vianello A. (2023). Relative Humidity and Its Impact on the Immune System and Infections. Int J Mol Sci. 24(11):9456.
- Harper M, John M, Turni C, Edmunds M, Michael FS, Adler B, et al. (2015). Rapid multiplex PCR assay for genotyping Pasteurella multocida strains using the lipopolysaccharide outer core biosynthesis locus. J Clin Microbiol. 53:477-485.
- García-Alvarez A, Vela AI, San Martín E, Chaves F, Fernández-Garayzábal JF, Lucas D, et al. (2017). Characterization of Pasteurella multocida associated with ovine pneumonia using multi-locus sequence typing (MLST) and virulence-associated gene profile analysis and comparison with porcine isolates. Vet Microbiol. 204:180-187.
- Wang J, Sang L, Sun S, Chen Y, Chen D, Xie X. (2019). Characterization of Pasteurella multocida isolated from dead rabbits with respiratory disease in Fujian, China. BMC Vet Res. 15(1):438.
- Hermans K, Devriese LA, Haesebrouck F. (2003). Rabbit staphylococcosis: difficult solutions for serious problems. Vet Microbiol. 91(1):57-64.
- Hecker M, Mäder U, Völker U. (2018). From the genome sequence via the proteome to cell physiology - Pathoproteomics and pathophysiology of Staphylococcus aureus. Int J Med Microbiol. 308(6):545-557.
- Schmidt T, Kock MM, Ehlers MM. (2017). Molecular Characterization of Staphylococcus aureus Isolated from Bovine Mastitis and Close Human Contacts in South African Dairy Herds: Genetic Diversity and Inter-Species Host Transmission. Front Microbiol. 8:511.
- Holliman A, Girvan GA. (1986). Staphylococcosis in a commercial rabbitry. Vet Rec. 119(8):187.
- Boucher HW, Corey GR. (2008). Epidemiology of methicillin-resistant Staphylococcus aureus. Clin Infect Dis. 46(Suppl 5):S344-S349.
- Silva V, De Sousa T, Gomez P, Sabenca C, Vieira-Pinto M, Capita R. (2020). Livestock-associated MRSA in purulent subcutaneous lesions of farm rabbits. Foods. 9(4):439.
- Dawod RE, Attya A, Arafat N. (2024). Staphylococcus aureus infection in rabbit farms: virulence genes and antimicrobial resistance. Egypt J Anim Health.
- Iorgoni V, Stanga L, Iancu I, Degi J, Popa I, Gligor A, et al. (2025). Multidrug-Resistant Escherichia coli Associated with Respiratory and Systemic Infection in a Domestic Rabbit in Romania: First Confirmed Case. Antibiotics. 14(9):929.
- Alam RTM, Fawzi EM, Alkhalf MI, Alansari WS, Aleya L, Abdel-Daim MM. (2018). Anti-Inflammatory, Immunomodulatory, and Antioxidant Activities of Allicin, Norfloxacin, or Their Combination against Pasteurella multocida Infection in Male New Zealand Rabbits. Oxid Med Cell Longev. 2018:1780956.
- El-Hendy AHM, Mohamed FM, Hassan AK. (2020). Clinicopathological and hematological changes following experimental infection of rabbits with Pasteurella multocida type A. SVU-IJVS. 3(2):1-13.
- El-Sheikh SMA, Youssef FM, Mohamed HI, El-Saber Batiha G, Albrakati A, Galal AAA. (2021). Grape seed hydro-alcoholic extract in treating rabbits experimentally infected with Pasteurella multocida. Vet Med Sci. 7(3):923-934.
- Peñailillo AK, Sepulveda MA, Palma CJ, Espinoza A, Aguilera M, Burgos RA, et al. (2016). Hematological and biochemical changes induced by low-dose Escherichia coli LPS in rabbits. Arch Med Vet. 48(3):315-320.
- Fatma MY. (1999). Clinicopathological studies on pasteurellosis in laboratory animals. MVSc Thesis, Suez Canal University.
- Ruble RP, Cullor JS, Brooks DL. (1999). Reactive thrombocytosis in New Zealand White rabbits in response to experimental Pasteurella multocida infection. Blood Cells Mol Dis. 25(2):95-102.
- Moustafa KME, El-Hosseiny HM, Shaheen GF, El-Kotamy EM, Ghoniem AE, Younan GE, et al. (2024). Impact of different forms of selenium supplementation on growth and physiological performance of New Zealand white rabbits. Trop Anim Health Prod. 56(4):131.
- Elmanylawi MF, El-Elaime R, Hekal SH, Mona F, Salama WA. (2025). Comparative growth response of New Zealand White rabbits to dietary selenium sources. Egypt J Vet Sci. pp. 1-6.
- Liu Y, Liu X, Xiang X, Qu X, Guo S. (2023). Effects of nano-selenium levels on growth, antioxidant capacity, biochemical parameters, and selenium content in Landes geese.
- Xia IF, Kong H-K, Wu MMH, Lu Y, Wong K-H, Kwok KWH. (2022). Selenium nanoparticles modulate immunity beyond redox effects: serum proteomics and transcriptomics. Antioxidants. 11(5):964.
- Bashar AM, Abdelnour SA, El-Darawany AA, Sheiha AM. (2022). Selenium nanoparticles and/or Spirulina platensis on growth, hematobiochemistry, antioxidant status, hormones, immunity, and apoptosis in heat-stressed rabbits. Egypt J Rabbit Sci. 32:77-103.
- Abdelmoteleb AMM, Mohamed FH, Zohree HA, Rezk RAA, Hamed EO, Elmahdy AM. (2023). Comparative impact of selenium and nano-selenium on cypermethrin-induced toxicity in rabbits. Egypt J Anim Health. 3(2):30-43.