Information Links
Related Conferences
Current Issue Volume 4, Issue 1 - 2025
Sensorineural Hearing Loss: Overview and E3 Ligase Inhibition for Transcription Factor ATOH1 Survival
Waseem Syed*
Medical Imaging, Lake Erie College of Osteopathic Medicine, Bradenton, USA
*Corresponding author: Mr. Waseem Syed, Medical Imaging, Lake Erie College of Osteopathic Medicine, Bradenton, 7507 Park Village Dr Jacksonville FL, 32256, USA, Tel: 4088056950; Email: [email protected]
Received Date: June 26, 2025
Published Date: July 23, 2025
Citation: Syed W. (2025). Sensorineural Hearing Loss: Overview and E3 Ligase Inhibition for Transcription Factor ATOH1 Survival. Mathews J Otolaryngol. 4(1):10.
Copyrights: : Syed W. © (2025).
ABSTRACT
This article is an examination of sensorineural hearing loss with potential reversal of symptoms through molecular modifications. Sensorineural hearing loss is a condition that plagues many patients, primarily the elderly, and can significantly lower the quality of life in these individuals. Although cochlear hair cells are incapable of regenerating, transcription factor atonal bHLH transcription factor 1 (ATOH1) is of interest as it is known to induce embryological differentiation of stem cells into hair cells. This article seeks to establish the basis of this transcription factor as a pharmacological target to aid those suffering from sensorineural hearing loss. Using data and information collected from previously published articles, manipulation of an organism’s natural protein recycling matrix, proteosome-ubiquitin system, via the use of E3 ligases may allow for prevention of the degradation of ATOH1 transcription factors. The compound is shown to have cysteine bases, which can be targeted utilizing electrophilic compounds. Results, however, have yet to be implemented in a laboratory environment but yield a promising opportunity for further evaluation of treating sensorineural hearing loss.
Keywords: Sensorineural Hearing Loss, Otolaryngology, ATOH1, Proteosome, E3 Ligases, Cystine Residues, Hearing Loss, Cochlea.
INTRODUCTION
The concept of molecular engineering for treatment of hearing loss is poorly studied. Before we can begin with a systematic review of how sensorineural hearing loss can be treated, it is worth spending some time describing the anatomy and embryology of the inner ear.
Sensorineural hearing loss, also known as SNHL, has two known etiologies. Some are of gradual onset occurring naturally as one ages or can be of sudden onset due to loud noises; Rarely, some people are born with it. It affects anywhere between one to six out of five thousand people every year. The primary causes of hearing loss are damage to either the hair cells in the cochlear membrane, or to the auditory portion of the vestibulocochlear nerve (cranial nerve eight). Of interest in this article is the damage done to the hair cells of the cochlear membrane [1].
The ear consists of three major structures, which include the outer ear, the middle ear, and the inner ear. The outer ear is concerned with conducting sound waves from the environment to the middle ear via vibration of the tympanic membrane, better known as the eardrum. The middle ear takes these vibrations and transmits them via three small bones, known as the malleus, incus, and stapes. The stapes articulates with the oval window where the sound waves are taken into the area of our primary interest, the inner ear. The inner ear houses many different organs, of which contain the vestibular organs for supporting balance (semicircular canals, utricle, saccule) and the cochlear organ for hearing [2].
The cochlea is the classic snail shaped organ where fluid moves in response to sound waves conducted by the middle ear (Figure 1). The basilar membrane inside houses the Organ of Corti [3] right on top of it. The organ has hair cells with stereocilia and kinocilium. Depolarization is marked by deflection of the stereocilia towards one direction, and hyperpolarization is marked by deflection of the stereocilia towards the opposite direction. The signal generated is transmitted via the auditory branch of cranial nerve eight (vestibulocochlear) [3] and synapses onto the cochlear nucleus in the brainstem, where the information is taken up to the primary auditory cortex in the temporal lobe to be processed (Figure 2).
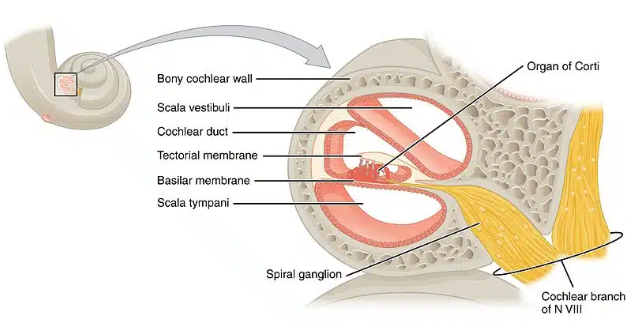
Figure 1. Anatomy of the inner ear cochlear organ, detailing the organ of corti
Image source: Davies et al., 2020 [3]; Used with permission from TeachMeAnatomy.
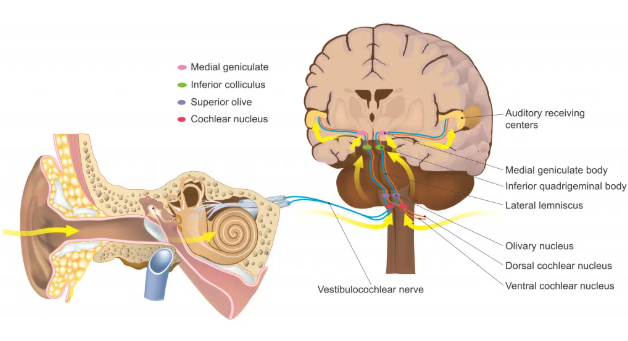
Figure 2. Signal pathway from the cochlear organ of the inner ear to the central nervous system.
Image source: Davies et al., 2020 [3]; Used with permission from TeachMeAnatomy.
Loud noise creates massive vibrations that causes the tiny hair-like projections on stereocilia to bend backwards with more intensity than lower noises. Over time, this damage builds up in subsequent stereocilia causing age related hearing loss [3]. While damage to the hair structures are permanent as they are not known to have any capability for regeneration, the embryogenesis of these cells yields some promising thoughts. Thus, it is worth mentioning the origin of the hair cells.
Embryogenesis
The notch pathway [4] has multiple roles for cochlear development. In short, the Notch pathway allows for basic cell-cell communication and controls proliferation and differentiation of many cells, hair cells included. This is done via the notch receptor cleavage from ligand activation, causing the release of Notch intracellular domain (NICD) which translocates to the nucleus where it interacts with DNA binding proteins. Overexpression of this pathway in mice have demonstrated patches of sensory hair cells. However, while it may have produced some promising results, there have been many other studies that have challenged this idea for hair cell regeneration. Thus, the aims for this article is better suited elsewhere, though it is worth mentioning the notch signaling pathway for hair cells.
The ATOH1 homolog is a basic helix-loop-helix transcription factor that gives rise to sensory hair cells, among others. Absence of this transcription factor caused hair cells to undergo apoptosis. Theoretically, as long as the hair cells can retain this transcription factor, they are able to regenerate. However, the Atoh-1 factor is eventually deleted at different developmental time points, limiting the regenerative capabilities of sensory hair cells. Thus, allowing for the stability of factor ATOH1 may be of primary interest, rather than said overexpression which other researchers have attempted to perform with little success. These results are highlighted in one of many experiments done by Izumikawa et al [5] where they highlight the adenovirus-based overexpression of ATOH1 transcription factor in non-sensory cells of flat epithelium. After many days, the flat epithelium did not demonstrate differentiation into sensory hair cells. Additionally, adding this therapy to cochlear cells did result in an increase of ATOH1, however there were no clear signs of hair cell regeneration. Abnormalities other than erratic projections of newly generated nerves after inducement with ATOH1were not detected. Additionally, testing in mice found no mortality from the previously mentioned study. While the action of inducing ATOH1 may be successful, the degradation of the transcription factor was not taken into account for, and thus may be a limiting factor in the study.
In essence, the focus on allowing for certain transcription factors to survive, namely ATOH1, may allow for sensory hair cells of the cochlea to remain intact and prevent the age-related or sudden decline of hearing seen in many patients worldwide.
Protein Degradation in the Cell
Although much more extensive research must be completed in order to fully understand ATOH-1 degradation, it is likely done via the ubiquitin-proteosome system, as this is how many transcription factors are degraded in the cells. Specifically, the 26s proteosome [6] is stationed in high concentrations in the nucleus that has the key function of degrading proteins in the nucleus, better known as transcription factors. To understand how we can protect ATOH-1, we must understand how this system works. Although simply deactivating the whole ubiquitin-proteosome system might seem plausible, this could yield devastating consequences as all transcription factors would be prevented from deletion, rather than just ATOH-1.
In essence, ubiquitin molecules, which are 8.6 kDa proteins are attached to lysine residues of a protein [7]. There is a 3 step process to this pathway that involves three separate enzymes: Ubiquitin activating enzyme (E1), Ubiquitin conjugating enzyme (E2), and Ubiquitin protein ligase (E3) [8]. E1 uses ATP molecules to activate the ubiquitination process where the ubiquitin molecule is transferred to E2. E3 is then used to transfer this ubiquitin onto a protein. This marks the protein for degradation, where complex is taken to the proteosome complex which contains a 20S core and a 19S complex with three active sites in the core (Figure 3). The marked proteins are then unfolded as they pass through the 20s core.
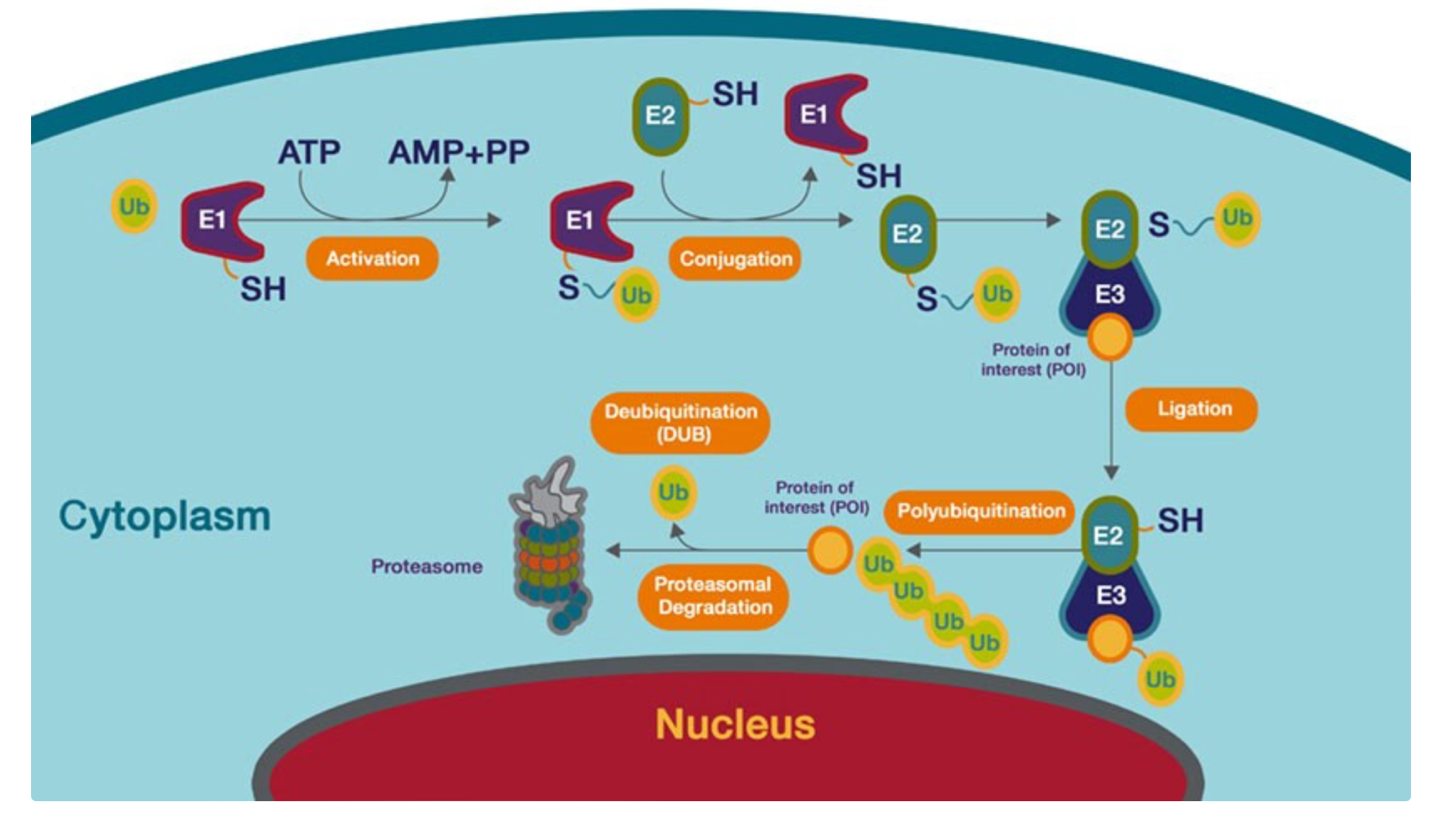
Figure 3. Diagram detailing the ubiquitination of lysine residues on a protein, marking it for degradation within the proteosome complex.
Image source: Thermo Fischer Scientific [7]; Used with permission from Thermo Fischer Scientific.
Prevention of Ubiquitination with E3 Ligases
E3 ligases have been heavily researched to both cause certain proteins to become degraded as well as inhibition of E3 ligases to prevent protein degradation via the proteosomes. Current study interests involve using PROTACs (proteolysis-targeting chimera) [9] to allow for degradation of certain proteins. Examples highlighted by the study performed by Lee J et al. [9] include potential inhibition of brain (FBXL16), pancreas (ASB9), and testis (DCAF4L1) E3 ligases to allow for treatment of certain diseases where overexpression of these ligases are seen. As seen in this study, the manipulation of E3 ligases can be a double-edged sword, where inhibiting the ligases can be used for keeping certain proteins alive while overexpression of E3 ligases can be utilized to degrade excessive production of proteins, as detailed in the previously mentioned study [9].
However, as mentioned before, E3 ubiquitin ligases may or may not be specific to each protein. This must be taken into consideration when inhibiting these ligases, as other proteins that need to be degraded can pile up and cause dysfunction. Other articles mention detecting specific E3 ligases substrates using the molecular technologies of CRISPER-Cas9 and shRNA screening. Identifying the specific E3 ligases that target ATOH-1 is what we hope to accomplish in the future, as inhibiting these will hopefully provide a better prognosis for hearing loss of sensorineural origin. While more research is being done, there are projects that involve base mutations of the zinc-binding region within RING domain of E3 ligase. This involves a cystine to histidine mutation [10].
Assessing Chirality of E3 Ligases
Assessing the chirality of E3 ligases is essential for future researchers to target these proteins for pharmacologic manipulation. There are four types of E3 ligases to consider: HECT type, U-box type, RING-finger type, and RBR type [11]. HECT ligase have the homologous to E6 associated protein C terminus that allows the activated E2 ligase to move ubiquitin proteins to an active cysteine site. Ring-finger E3 ligases have a RING domain where the ubiquitin protein is actually transferred from E2 directly to the substrate, bypassing the need for the R3 intermediate. U-box E3 ligases are known for controlling the quality of proteins in cells. The structure of the U-box is similar to the RING domain in that it is needed to accept the substrate from E2 ligases. RBR E3 ligases are unique in that they have two regions. They have RING1 which is used to interact with E2 ligases and RING2 which has the catalytic cystine. All of these different types of E3 ligases are depicted with their basic structure in the image below (Figure 4). Through these different types of E3 ligases, we can understand that the catalytic cystine is necessary for some, if not all, of the E3 ligases. Pharmacologic manipulation of these active site can yield these ligases to become obsolete.
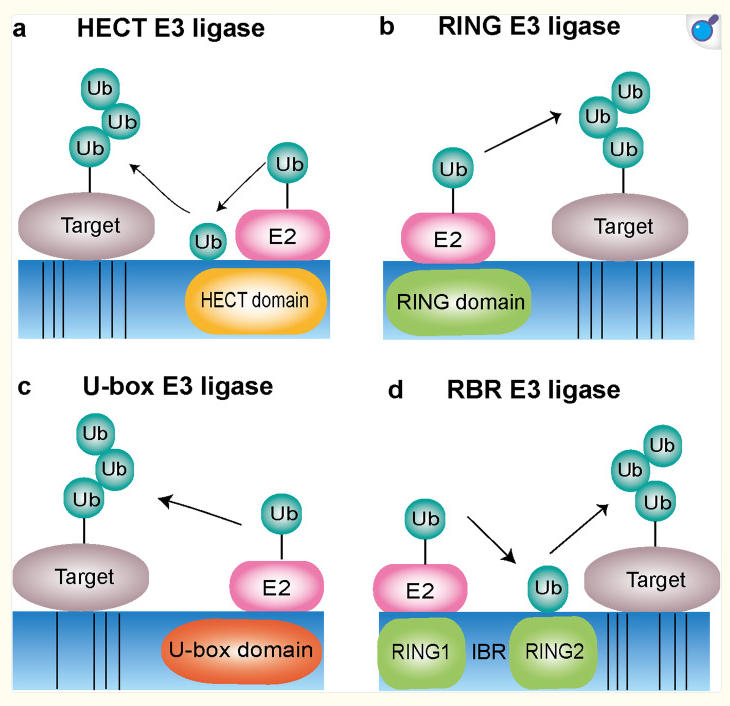
Figure 4. Diagram depicting the four different types of E3 ligases: HECT E3 ligase, RING E3 ligase, U-box E3 ligase, RBR E3 ligase.
Image Source: Yang Q et al., 2021 [11]; Used with permission from National Library of Medicine.
Pharmacological Manipulation
Cystine is an amino acid known for its sulfur component, which is the basis of formation of disulfide bonds (Figure 5).
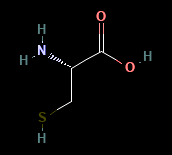
Figure 5. Molecular compound of the amino acid Cystine depicting its individual atoms.
Image source: National Center for Biotechnology Information, 2025 [12]; Used with permission from Biotechnology Information.
To render the cystine molecular obsolete, many different compounds can be used. In particular, using the thiol group (S-H) which is unique to cystine, electrophilic compounds can attack the heavily neutrophilic structure of the thiol group. Some commonly used electrophiles are epoxide compounds and acrylamide. Both are heavily electron dense and can attack the electron weak areas of the thiol bond. By targeting these cysteine sites in E3 ligases using electrophilic compounds, we can render the ubiquitination process of proteins useless. More research is needed to implement and test these theories.
CONCLUSION
Sensorineural hearing loss is a condition that plagues many individuals, ranging in ages (young and elderly) and speed of onset (sudden or gradual). There are two main causes, of which this article focused on the damage done to hair cells in organ of Corti. ATOH1 transcription factor is expressed in early development of sensory hair cells but is later degraded and is found in low concentrations in mature cells. The primary mechanism for this is via degradation of proteins in the nucleus by marking it with ubiquitination, causing the destruction of the marked protein in the proteosome. E3 ligases are the final step in the three-step pathway for marking a protein for destruction. The four different types of E3 ligases all share the same factor in that they have an active cysteine site which is crucial for their function. Cystine contains a sulfur based thiol group which can be attacked by electrophilic compounds, rendering the cystine amino acid useless and thus rendering the ubiquitination process obsolete. Further research is needed for implementation and testing. This will hopefully provide a better treatment option for those experiencing hearing loss of sensorineural origin.
ACKNOWLEDGEMENTS
None.
CONFLICTS OF INTERESTS
There are no conflicts of interests.
REFERENCES
- Sensorineural Hearing Loss (SNHL): What You Should Know. Cleveland Clinic. February 21, 2025. Available at: https://my.clevelandclinic.org/health/diseases/sensorineural-hearing-loss
- What Is the Inner Ear? Cleveland Clinic. August 29, 2024. Available at: https://my.clevelandclinic.org/health/body/24340-inner-ear
- Davies, Kristen, and Yuri Sugano. The Auditory Pathway. TeachMeAnatomy. December 14, 2020. Available at: https://teachmeanatomy.info/neuroanatomy/pathways/auditory-pathway/
- Atkinson PJ, Najarro EH, Sayyid ZN, Cheng AG. (2015). Sensory Hair Cell Development and Regeneration: Similarities and Differences. Development. 142 (9):1561-1571.
- Izumikawa M, Batts SA, Miyazawa T, Swiderski DL, Raphael Y. (2008). Response of the flat cochlear epithelium to forced expression of Atoh1. Hear Res. 240(1-2):52-56.
- Muratani M, Tansey WP. (2003). How the Ubiquitin–Proteasome System Controls Transcription. Nat Rev Mol Cell Biol. 4(3):192-201.
- Protein Degradation Using the Ubiquitin-Proteasome Pathway. Thermo Fisher Scientific – US. Available at: www.thermofisher.com/us/en/home/life-science/cell-analysis/cell-analysis-learning-center/protein-degradation-resource-center/protein-degradation-using-ubiquitin-proteasome-pathway.html
- Guo HJ, Rahimi N, Tadi P. (2025). Biochemistry, Ubiquitination. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK556052/
- Lee J, Lee Y, Jung YM, Park JH, Yoo HS, Park J. (2022). Discovery of E3 Ligase Ligands for Target Protein Degradation. Molecules. 27(19):6515.
- Lydeard J, Harper J. (2010). Inhibitors for E3 ubiquitin ligases. Nat Biotechnol. 28(7):682-684.
- Yang Q, Zhao J, Chen D, Wang Y. (2021). E3 ubiquitin ligases: styles, structures and functions. Mol Biomed. 2(1):23.
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5862, Cysteine. PubChem. Available at: https://pubchem.ncbi.nlm.nih.gov/compound/Cysteine.