Previous Issues Volume 1, Issue 1 - 2016
Perturbing Transsulfuration Pathway and Sulfur Recycling, A New Direction Towards Therapeutic Intervention of Inflammatory Bowel Disease
Sanjib Bhattacharyya1
Corresponding Author: Sanjib Bhattacharyya, WPI-Advanced Institute for Materials Research (WPI-AIMR), Tohoku University, Sendai, Japan. Tel: +81-75-753-9864; E-mail:[email protected]
Received Date: 28 Mar 2016 Accepted Date: 05 May 2016 Published Date: 07 May 2016
Copyright © 2016 Bhattacharyya S
Citation:Bhattacharyya S. (2016). Perturbing Transsulfuration Pathway and Sulfur Recycling, A New Direction Towards Therapeutic Intervention of Inflammatory Bowel Disease. Mathews J Gastroenterol Hepatol 1(1): 001.
ABSTRACT
Combating inflammatory bowel disease (IBD) is becoming a growing challenge to the global health management. Our bowel can be envisioned as a big vessel reactor where millions of reactions occur at each moment, every day to meet the physiological demand of our body for quality control of our life. This includes the orchestration of several steps which ultimately funnel down to the intestinal system to drives the end point of the digestion process. Intestinal sulfur recycling by transsulfuration pathway can modulates the integrity of our bowel symptoms. Limiting the sulfur amino acid metabolism and mitigating hydrogen sulfide (H2S) generation can help to stabilize the colonic biochemistry with monitoring of metabolites for subsequent health risk assessment. Regulating protein sulfhydration in colonic tissue could play detrimental roles in the modulation of several contributing factors related to IBD.
KEYWORDS
Transsulfuration pathway; Hydrogen sulphide; Sulfhydration; Inflammatory bowel disease; Biomarkers; Therapy.
INTRODUCTION
BACKGROUND
Factors Related to IBD Pathogenesis Connecting Transsulfuration Homeostasis:
Environment:Our bowel is challenged daily with a load of foods, primarily sourced from the environment. Many food components contain sulfides, polysulfide compounds, which ultimately breakdown to generate gaseous H2S in situ [1, 2]. Hydrogen sulfide can interact with metabolic cytochrome c oxidase and carbonic anhydrase, by chelating with metalbinding porphyrin cofactor [3]. Evolutionarily, H2S has been viewed as an important prebiotic element [4]. In nature, H2S concentration increases in the deep ocean, oceanic volcanoes and mountain spring while the oxygen concentration decreases. Importantly, hydrogen sulfide can bypass the cellular oxygen demand and modulate energy production [5]. Thus, H2S can sense the hypoxia related changes inside the cell [6]. Micro biome: There are many micro bacteria that can produce H2S [7]. Some of these bacteria exist in the gut mucosal system of our gastrointestinal tract [8, 9]. Examples of these bacteria include sulfate reducing bacteria (SRB), thiobacillus, desulfobacter among others. Some of this microbiota could play a potent role during GI infection and inflammation, and thus could be one of the leading causes of IBD [8, 10-13]. Therefore, H2S producing colonic microbiota in the GI tract can also modulate the mucosal immune system, and the IBD state, by producing H2S or sulfur metabolites [8, 11, 13]. Immune Response: Immune surveillance in the GI tract plays a determining role in the progression of IBD [9, 14]. Our mucosal immune checkpoint does its best by T cell activation and adaptive immunities, like dendritic cell, and macrophage activation to nullify the toxins components that enter in our bowl during food intake [15]. It stimulates the cytokine (TNF-?, IFN-?) and chemokine production by the processing of the MHC class I & II peptides [10, 11]. H2S can trigger T cell activation [16]. Mammalian tissue contains enzymes, Cystathionine beta synthase (CBS) and Cystathionine gamma lyase (CSE) that can endogenously produce H2S (Figure 1) [3]. H2S has been implicated in many inflammatory disease conditions, such as pancreatitis, sepsis, and joint and pulmonary inflammation [3, 17-19].
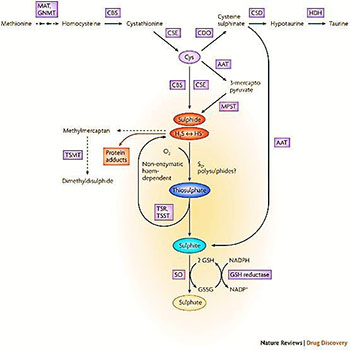
Figure1: Enzymatic pathways of H2S production in mammalian cells.
Methionine, which is derived from alimentary sources, is converted to S-adenosylmethionine by methionine adenosyltransferase (MAT). S-adenosylmethionine is subsequently hydrolysed to homocysteine by glycine N-methyltransferase (GNMT). Cystathionine-synthase (CBS) catalyses the production of cystathionine by transferring serine to homocysteine. Cystathionine-lyase (CSE), a pyridoxal 5’-phosphate-dependent enzyme, subsequently converts cystathionine to cysteine (Cys). CSE catalyses a -disulphide elimination reaction that results in the production of pyruvate, NH4+ and thio cysteine. Thiol cysteine may react with cysteine or other thiols to form hydrogen sulphide (H2S). One pathway of cysteine metabolism involves its oxidation to cysteine sulphinate by cysteine deoxygenase (CDO), which then gets further converted to hypotaurine by cysteine sulphinate decarboxylase (CSD), and subsequently to taurine by a nonenzymatic reaction or by hypotaurine dehydrogenase (HDH). The above reactions predominantly take place in the cytosol. In the mitochondria, cysteine can get converted to 3-mercaptopyruvate by aspartate aminotransferase (AAT), which can then be converted to H2S by 3-mercaptopyruvate sulphur transferase (MPST). Sulphide, via non-enzymatic reactions, gets metabolized to thiosulphate (one molecule of sulphide yields two molecules of thiosulphate), which then gets converted to sulphite by thiosulphate reductase (TSR), for instance in liver, kidney or brain tissues or thiosulphate sulphurtransferase (TSST), which is predominantly expressed in the liver. The conversion of cysteine sulphinate to sulphinyl pyruvate by AAT, followed by a non-enzymatic reaction, can also yield sulphite. Sulphite gets oxidized to sulphate by sulphite oxidase (SO) by a glutathione (GSH)-dependent process. H2S can also yield protein adducts, and can be converted to methylmercaptan and dimethyldisulphide by thiol-S-methyltransferase (TSMT). Non-enzymatic oxidation of sulphide can also yield the generation of polysulphides and elemental sulphur (S0). Reprinted with permission from Ref (3), Csaba Szabo. Hydrogen sulfide and its therapeutic potential; Nature review drug discovery. 2007, 6 (11), 917-935. Copyright©2016 Nature Publishing Group (npg). Genome:Given the recent understanding on how several genomic alterations play a key role in the pathophysiological phenotype of many diseases, more targeted therapy can be achieved in IBD. The metabolic burden could trigger unfolded protein response (UPR) and thus (endoplasmic reticulum) ER stress to the colonic tissue. This event could lead to the upregulation (EIF2A, ATF6, IRE1, and PERK) and down regulation of several genes [10, 20]. H2S can also cause ER stress by protein sulfhydration of protein tyrosine phosphatase 1B (PTP1B), a newly identified post translational modification [21]. Additionally, homocysteine, a component of sulfur amino acid metabolism, is also known to trigger ER stress [22]. Homocysteine is found to be high in fistula patients and possibly modulates methionine dependent genomic alteration [23, 24]. H2S mediated sulfhydration of transcription factor (NF-kB) leads to the induction of anti-apoptotic genes [25, 26]. H2S can even inhibit the phenyl isothiocyanate (cancer preventive) mediated apoptosis of colon cancer [27]. Recent reports also show that CBS plays a pivotal role in the chemo resistance of various cancers [28, 29]. This raises the possibility that the antimicrobial agents that initially cure the gut mucosal inflammation during IBD, might develop resistance, possibly by sulfur amino acid metabolite.
Maturation of IBD by Transsulfuration and Sulfide Homeostasis: Hydrogen sulfide can activate the macrophages and T cell to produce the pro-inflammatory cytokines (TNF-ɑ,IL-1β, IL-6) [16, 19]. The gut mucosal bacteria can also produce harmful chemokine and cytokines after activation [8]. Thus, it implicates that over the progression of IBD, H2S acts as an pro-inflammatory molecule (scheme 1) [30]. At advanced stages of the disease, endogenous formation of H2S could be augmented due to increase of CBS transcriptome, as observed in colon cancer [31]. In pathological condition, excess sulfide could be further pumped out by gut mucosal bacteria to promote H2S production [13, 32, 33]. H2S can induce ER stress by NF-kB activation and protein sulfhydration during reductive stress to adjust the redox homeostasis (scheme 1) [25, 26]. Hydrogen sulfide modified NF-kB leads to the anti-apoptotic effect of the cell by protein sulfhydration. The protein sulfhydration by H2S might impact many important cellular events, such as autophagy, lipophagy and other quality control genomic loci, including H2S synthesizing genes (CBS) [20, 34]. Additionally, the upregulation of homocysteine can cause ER stress and alteration of sterol-triglyceride synthesizing genes [22]. H2S mediated sulfhydration and sulfation of targets proteins possibly cause the chemo attraction of immune cells [35]. Beside H2S, it's metabolites (thiosulfate, sulfite, and sulfate), could also modify protein (tyrosine sulfation) and initiate the immune cell infiltration to trigger inflammation [35, 36]. However, sulfhydration or sulfation guided prolonged infiltration of immune cell to the colonic tissue, could direct the IBD towards an auto immune disorder. Cellular hypoxia can also facilitate the mucosal colonic damage by altering oxygen gradient and metabolic junctions, thus trigger inflammation to the bowl.37 Importantly, It has been reported that hydrogen sulfide can bypass the metabolic oxygen demand and modulate energy production at mitochondria [5, 29, 31]. Being a pre-biotic element and same family member of periodic table (group VI/16), it is possible that nature has evolutionarily preserved the sulfide generation either to mimic the oxygen demand during cellular hypoxia (scheme 1). Therefore, under hypoxic condition, the cellular environment can shift towards augmented production of H2S and H2S producing enzymes CBS to aggravate the severity of IBD. In addition, the microbiota can produce H2S to overcome mucosal barrier in the microenvironment leading to IBD progression. Suppressing H2S production could prevent excess sulfhydration mediated post-translational modification of protein. Inhibition of CBS could also impact on post genomic modulation of DNA methylation landscape (scheme 1) during IBD (such as hypo or hyper methylation) as observed in other chronic liver disease [38, 39]. An imbalance in sulfur homeostasis could be pivotal in the advancement of IDB. Hence, CBS inhibition can diminish the H2S synthesis and many other sulfur metabolite related alternation of cell signaling events, thereby preventing the aggressive onset of IBD.
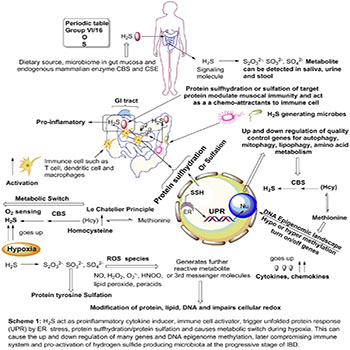
Scheme 1: Novel treatment strategy for IBD management by targeting CBS;
Role of hydrogen sulfide and its metabolite, protein sulfhydration and endogenous H2S producing machinery (CBS/CSE) in advancement of IBD and possible and non-invasive bio-marker for health risk assessment for IBD. Upregulation of transsulfuration and hydrogen sulfide generation can cause excess protein sulfhydration that can causes several cascades of events such as immune modulation, oxygen sensing in hypoxic tissue, ER stress as UPR and alteration of pro-inflammatory cytokine-chemokine level. Imbalanced in transsulfuration can also change the epigenome by pushing the equilibrium toward the augmented production of methionine. Generation of reactive sulfur species by combination with other reactive oxygen species can modify the DNA, protein and lipid to impair cellular redox.
Technological Advances to Monitor Transsulfuration for IBD Assessment: Hydrogen sulfide is a highly water and lipid soluble gaseous compound. In case of H2S poisoning, H2S and its metabolite S2O3 2- can be found in high concentration in urine and blood/ serum, and treatment of hydroxocobalamin can be used for detoxification [40, 41]. H2S can be detected in urine and stool [42]. Commercially developed analytical electrodes can measure H2S in various biological and preclinical samples such as blood, serum, and urine at low concentration in the order of 100 nM [43]. Besides, there are several other method such as HPLC, colorimetry, amperometry, voltammetry, titrimetric, fluorometry, turbid metric, led acetate trap analysis exits that is routinely used. Specially, the measurement of H2S metabolite (thiosulfate, sulfate and sulfite), as used previously by forensic department during sulfide poisoning [40, 44-46]. Given recent advances in imaging, H2S can be imaged in living mammals to locate the actuate state of inflammation in accordance with sulfide production [47]. A dye Sulfidefluor-5 probe (SF5) that specifically lit up by H2S synthesis in vivo, has been used to image the mice during zymosan mediated inflammation triggered by immune filtration (Figure 2) [47]. In addition to this study, a chemo luminescent dye Spiro Adamantane 1,2-dioxetane has been recently reported that can lit up by H2S generation, both in vitro and in vivo [48]. This dye can image H2S production both in solution (buffer) as well as in mice when treated with Na2S (in situ H2S generator), compared to a vehicle control. This dye can be further used for the evaluation of several pre-clinical animal model of IBD to confirm whether the phenotypic onset of IDB is related to the endogenous production of hydrogen sulfide or not. This will further implicate the physiological role of H2S during IBD progression, and thereby the regulation of sulfur amino acid metabolism for the development of next generation therapeutics for IBD management. Besides imaging, bacteriology profile of the sulfur regulating bacteria (such as sulfate reducing bacteria (SRB), thiobacillus, desulfobacter, etc.) in feces and urine sample could be analyzed as the early prognostic marker of IBD [42, 49-51]. The micro biome culture for H2S and its metabolites in the urine and feces samples might tell us the severity of the IBD [13, 42].
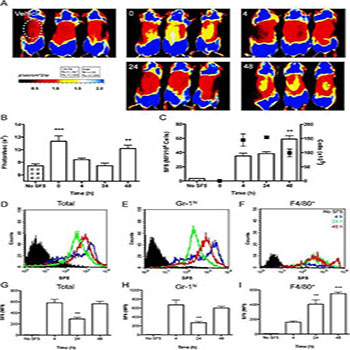
Figure 2: Acute inflammation impairs both tissue and cellular hydrogen sulfide synthesis.
Tissue and cellular incorporation of SF5 probe was measured over 48 h in a model of zymosan-induced air pouch inflammation; data are representative of two experiments done in triplicate. Vehicle or SF5 (100 μl; 10 μM)were injected into the pouch 30 min prior to whole mouse imaging with a IVIS Spectrum (Caliper Life Sciences) during a 48 h time course (A). Fluorescence emission was measured within a region of interest (ROI; white dotted line) encapsulating the air pouch expressed as photons/sec over the time course (B). SF5 fluorescence intensity in inflammatory exudates was analyzed by spectrofluorometry (C) and expressed as a ratio of infiltrating cell number (n = 3). Total cell numbers are overlaid (black points; right Y-axis). Reprinted with permission from Ref (47). Neil. Dufton, Jane. Natividad, Elena F. Verdu, and John L. Wallace. Hydrogen sulfide and resolution of acute inflammation: A comparative study utilizing a novel fluorescent probe, Scientific Report, 2012, 2, 499. Copyright© 2016 Nature Publishing Group (npg).
CONCLUSION
Regulation of transsulfuration pathway by inhibiting the cystathionine beta synthase (CBS) enzyme could be a potential target for the treatment of IBD. Targeted therapy by using small molecule inhibitors and gene silencing of CBS could be helpful to prevent the severe onset of IBD. Sulfur amino acid metabolite (hydrogen sulfide, thiosulfate, sulfite, sulfate and homocysteine) in the saliva, as well as in urine, feces could give us predictive warnings for the non-invasive health risk assessment of IBD (scheme 1). Several methods can be adopted for the targeted drug delivery of small molecule inhibitors and gene silencer (siRNA) specific to CBS enzyme. For examples, siRNA or anti-sense oligonucleotide for CBS gene can be used with nano liposomal formulation [29]. Similarly, other formulation like PLGA-PEG microsphere can be used to formulate the CBS siRNA or small molecule inhibitor of CBS. Given only few numbers of CBS specific inhibitors exists, it will be important to develop library of compounds by high throughput screening to come up with a potent and selective CBS specific inhibitors. In addition to these, many other polymer capsule strategies that are heavily used in the pharmaceutical industry can be adopted to prepare small molecule oral drugs formulations for the prevention of IBD. Selective inhibition of CBS with differential regulation of hydrogen sulfide activity remains an open challenge to scientific community for future management of IBD.
REFERENCES
- Benavides GA, Squadrito GL, Mills RW, Patel HD, et al. (2007). Hydrogen sulfide mediates the vasoactivity of garlic. Proc Natl Acad Sci, USA. 104(46), 17977-17982.
- Magee EA, Richardson CJ, Hughes R and Cummings JH. (2000). Contribution of dietary protein to sulfide production in the large intestine: an in vitro and a controlled feeding study in humans. The American Journal of Clinical Nutrition. 72(6), 1488-1494.
- Szabo C. (2007). Hydrogen sulphide and its therapeutic potential. Nat Rev Drug Discov. 6(11), 917-935.
- Parker ET, Cleavesb HJ, Dworkinc JP, Glavin DP, et al. (2011). Primordial synthesis of amines and amino acids in a 1958 Miller H2S-rich spark discharge experiment. Proceedings of the National Academy of Sciences.
- Fu M, Zhang W, Wu L, Yang G, et al. (2012). Hydrogen sulfide (H2S) metabolism in mitochondria and its regulatory role in energy production. Proceedings of the National Academy of Sciences. 109(8), 2943-2948.
- Olson KR. (2013). Hydrogen sulfide as an oxygen sensor. Clin Chem Lab Med. 51(3), 623-632.
- Lloy D. (2006). Hydrogen sulfide: clandestine microbial messenger? Trends in microbiology. 14(10), 456-462.
- Carbonero F, Benefiel AC, Alizadeh-Ghamsari AH and Gaskins HR. (2012). Microbial pathways in colonic sulfur metabolism and links with health and disease. Front Physiol. 3, 448.
- Chen W, Liu F, Ling Z, Tong X, et al. (2012). Human intestinal lumen and mucosa-associated microbiota in patients with colorectal cancer. PLoS One. 7(6), e39743.
- Cho JH. (2008). The genetics and immunopathogenesis of inflammatory bowel disease. Nat Rev Immunol. 8(6), 458-466.
- Arpaia N, Campbell C, Fan X, Dikiy S, et al. (2013). Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature. 504(7480), 451-455.
- Furusawa Y, Obata Y, Fukuda S, Endo TA, et al. (2013). Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature. 504(7480), 446-450.
- Pitcher MC and Cummings JH. (1996). Hydrogen sulphide: a bacterial toxin in ulcerative colitis? Gut. 39(1), 1-4.
- Kaser A, Zeissig S and Blumberg RS. (2010). Inflammatory Bowel Disease. Annual Review of Immunology. 28, 573-621.
- Hur SJ, Kang SH, Jung HS, Kim SC, et al. (2012). Review of natural products actions on cytokines in inflammatory bowel disease. Nutr Res. 32(11), 801-816.
- Miller TW, Wang EA, Gould S, Stein EV, et al. (2012). Hydrogen sulfide is an endogenous potentiator of T cell activation. J Biol Chem. 287(6), 4211-4221.
- Whiteman M and Winyard PG. (2011). Hydrogen sulfide and inflammation: the good, the bad, the ugly and the promising. Expert Rev Clin Pharmacol. 4(1), 13-32.
- Bhatia M. (2012). Role of Hydrogen Sulfide in the Pathology of Inflammation. Scientifica 2012, 12.
- Zhi L, Ang AD, Zhang H, Moore PK, et al. (2007). Hydrogen sulfide induces the synthesis of proinflammatory cytokines in human monocyte cell line U937 via the ERK-NF-kappaB pathway. J Leukoc Biol. 81(5), 1322-1332.
- Bogaert S, De Vos M, Olievier K, Peeters H, et al. (2011). Involvement of endoplasmic reticulum stress in inflammatory bowel disease: a different implication for colonic and ileal disease? PLoS One. 6(10), e25589.
- Krishnan N, Fu C, Pappin DJ and Tonks NK. (2011). H2S-Induced sulfhydration of the phosphatase PTP1B and its role in the endoplasmic reticulum stress response. Sci Signal. 4(203), ra86.
- Werstuck GH, Lentz SR, Dayal S, Hossain GS, et al. (2001). Homocysteine-induced endoplasmic reticulum stress causes dysregulation of the cholesterol and triglyceride biosynthetic pathways. J Clin Invest. 107(10), 1263-1273.
- Kelly D. (1999). Nutrition in inflammatory bowel disease. Current Gastroenterology Reports. 1(4), 324-330.
- Mallamaci F, Bonanno G, Seminara G, Rapisarda F, et al. (2005). Hyperhomocysteinemia and arteriovenous fistula thrombosis in hemodialysis patients. Am J Kidney Dis. 45(4), 702-707.
- Sen N, Paul BD, Gadalla MM, Mustafa AK, et al. (2012). Hydrogen sulfide-linked sulfhydration of NF-kappaB mediates its antiapoptotic actions. Mol Cell. 45(1), 13-24.
- Bindu DP and Solomon HS. (2012). H2S signalling through protein sulfhydration and beyond. Nature Reviews Molecular Cell Biology. 13, 499-507.
- Rose P, Moore PK, Ming SH, Nam OC, et al. (2005). Hydrogen sulfide protects colon cancer cells from chemopreventative agent beta-phenylethyl isothiocyanate induced apoptosis. World J Gastroenterol. 11(26), 3990-3997.
- Ryu CS, Kwak HC, Lee KS, Kang KW, et al. (2011). Sulfur amino acid metabolism in doxorubicin-resistant breast cancer cells. Toxicol Appl Pharmacol. 255(1), 94-102.
- Bhattacharyya S, Saha S, Giri K, Lanza IR, et al. (2013). Cystathionine Beta-Synthase (CBS) Contributes to Advanced Ovarian Cancer Progression and Drug Resistance. PLoS One. 8, e79167.
- Seril DN, Liao J, Yang GY and Yang CS. (2003). Oxidative stress and ulcerative colitis-associated carcinogenesis: studies in humans and animal models. Carcinogenesis. 24(3), 353- 362.
- Szabo C, Coletta C, Chao C, Módis K, et al. (2013). Tumorderived hydrogen sulfide, produced by cystathionine-betasynthase, stimulates bioenergetics, cell proliferation, and angiogenesis in colon cancer. Proc Natl Acad Sci, USA. 110(30), 12474-12479.
- Ng Y-L, Klopcic B, Lloyd F, Forrest C, et al. (2013). Secreted Protein Acidic and Rich in Cysteine (SPARC) Exacerbates Colonic Inflammatory Symptoms in Dextran Sodium Sulphate- Induced Murine Colitis. PLoS One. 8, e77575.
- Medani M, Collins D, Docherty NG, Baird AW, et al. (2011). Emerging role of hydrogen sulfide in colonic physiology and pathophysiology. Inflamm Bowel Dis. 17(7), 1620-1625.
- Hetz C, Chevet E and Harding HP. (2013). Targeting the unfolded protein response in disease. Nat Rev Drug Discov. 12, 703-719.
- Al-Ubaidi MR and Kannan Y. (2013). Tyrosine O Sulfation: An Overview. JSM Biotechnol Bioeng. 1(1), 5.
- Moore KL. (2009). Protein tyrosine sulfation: a critical posttranslation modification in plants and animals. Proc Natl Acad Sci, USA. 106(35), 14741-14742.
- Colgan SP and Taylor CT. (2010). Hypoxia: an alarm signal during intestinal inflammation. Nat Rev Gastroenterol Hepatol. 7(5), 281-287.
- Mato JM, Martinez-Chantar ML and Lu SC. (2008). Methionine metabolism and liver disease. Annu Rev Nutr. 28, 273- 293.
- Finkelstein JD. (2003). Methionine metabolism in liver diseases. Am J Clin Nutr. 77, 1094-1095.
- Fujita Y, Fujino Y, Onodera M, Kikuchi S, et al. (2011). A fatal case of acute hydrogen sulfide poisoning caused by hydrogen sulfide: hydroxocobalamin therapy for acute hydrogen sulfide poisoning. J Anal Toxicol. 35(2), 119-123.
- Levitt MD, Furne J, Springfield J, Suarez F, et al. (1999). Detoxification of hydrogen sulfide and methanethiol in the cecal mucosa. J Clin Invest. 104, 1107-1114.
- Levine J, Ellis CJ, Furne JK, Springfield, et al. (1998). Fecal hydrogen sulfide production in ulcerative colitis. Am J Gastroenterol. 93(1), 83-87.
- Yang G, Wu L, Jiang B, Yang W, et al. (2008). H2S as a Physiologic Vasorelaxant: Hypertension in Mice with Deletion of Cystathionine ?-Lyase. Science. 322(5901), 587-590.
- Beauchamp RO, Bus JS, Popp JA, Boreiko CJ, et al. (1984). A critical review of the literature on hydrogen sulfide toxicity. Crit Rev Toxicol. 13(1), 25-97.
- Durand M and Weinstein P. (2007). Thiosulfate in human urine following minor exposure to hydrogen sulfide: implications for forensic analysis of poisoning. Forensic Toxicology. 25(2), 92-95.
- Snyder JW, Safir EF, Summerville GP and Middleberg RA. (1995). Occupational fatality and persistent neurological sequelae after mass exposure to hydrogen sulfide. Am J Emerg Med. 13(2), 199-203.
- Dufton N, Natividad J, Verdu EF and Wallace JL. (2012). Hydrogen sulfide and resolution of acute inflammation: A comparative study utilizing a novel fluorescent probe. Sci Rep. 2, 499.
- Cao J, Lopez R, Thacker JM, Moon JY, et al. (2014). Chemiluminescent probes for imaging H2S in living animals. Chemical Science. 6, 1979-1985.
- Fite A, Macfarlane GT, Cummings JH, Hopkins MJ, et al. (2004). Identification and quantitation of mucosal and faecal desulfovibrios using real time polymerase chain reaction. Gut. 53(4), 523-529.
- Zinkevich VV and Beech IB. (2000). Screening of sulfatereducing bacteria in colonoscopy samples from healthy and colitic human gut mucosa. FEMS Microbiol Ecol. 34(2), 147- 155.
- Gibson GR, Macfarlane GT and Cummings JH. (1988). Occurrence of sulphate-reducing bacteria in human faeces and the relationship of dissimilatory sulphate reduction to methanogenesis in the large gut. J Appl Bacteriol. 65(2), 103-111.